Пути специальной чувствительности
Рассмотрение путей специальной чувствительности целесообразно провести с позиций филогенеза, по мере формирования нервных центров в головном мозге. Центры специальной чувствительности развивались в следующем порядке: вначале центр равновесия, затем слуха, зрения, вкуса и, наконец, обоняния. Указанные центры связаны с соответствующими афферентными путями и органами, в которых располагаются рецепторы. Морфофункциональная связь таких нервных структур, как рецепторы, афферентные проводящие пути и связанные с ними центры, представляет собой анализатор определенных видов чувствительности.
1. Слуховой анализатор обеспечивает восприятие звуковых раздражений, проведение нервных импульсов до слуховых нервных центров, анализ и интеграцию поступившей в них информации. Функция слухового анализатора многоплановая. Прежде всего, слуховой анализатор создает возможность общения между людьми с помощью речи. Слух позволяет получать и анализировать звуковую информацию, поступающую из внешней среды, определять направление звука, его силу, тембр. Благодаря слуховой памяти мы можем определять принадлежность звука определенному человеку или предметам.
Рецепторы, воспринимающие звуковые раздражения, располагаются в органе слуха (кортиевом органе). Кортиев орган находится в улитковом протоке и представлен волосковыми клетками.
Механизм восприятия звука достаточно сложен. Вначале звук улавливается ушной раковиной, направляется в наружный слуховой проход и вызывает колебания барабанной перепонки. Ее колебания передаются на слуховые косточки – молоточек, наковальню и стремечко. Последнее своим основанием закрывает овальное окно преддверия лабиринта и вызывает перемещение перилимфы по лестнице преддверия, а затем – барабанной лестнице. Колебания перилимфы передаются эндолимфе вторичной барабанной перепонкой. Колебания базилярной пластинки, на которой находится кортиев орган, совершаются не на всем ее протяжении, а только в тех участках, которые резонансно совпадают с частотой звуковой волны. Установлено, что для низких звуков такие участки мембраны находятся у вершины улитки, для высоких – у основания. Ухо человека воспринимает звуковые волны с частотой колебаний от 16 до 21 тыс. Гц. Для речевых звуков оптимальный интервал составляет 1000–4000 Гц. Восприятие звука происходит за счет раздражения волосковых клеток, находящихся на определенном участке базилярной мембраны. Механическое раздражение трансформируется в нервный импульс, который передается с волосковых клеток на периферические отростки биполярных клеток.
Тела биполярных клеток (первый нейрон) образуют улитковый (спиральный) узел, расположенный в спиральном канале улитки внутреннего уха. Центральные отростки биполярных нейронов собираются в пучок, который называют улитковым корешком преддверно-улиткового нерва. Данный корешок проходит через отверстие на дне внутреннего слухового прохода, присоединяется к преддверному корешку и направляется к мосто-мозжечковому углу, где вступает в вещество моста и заканчивается на клетках переднего и заднего улитковых ядер (см. рис. 3.8). Эти ядра располагаются в латеральном углу ромбовидной ямки и содержат вторые нейроны слухового пути.
Аксоны клеток переднего улиткового ядра направляются в медиальном направлении и заканчиваются преимущественно на нейронах переднего ядра трапециевидного тела противоположной стороны. Аксоны клеток заднего улиткового ядра выходят на дорсальную поверхность моста в виде мозговых полосок и пересекают ромбовидную ямку в поперечном направлении. В области срединной борозды они погружаются в вещество мозга и в составе трапециевидного тела достигают его заднего ядра противоположной стороны. Таким образом, третьи нейроны слухового пути располагаются в ядрах трапециевидного тела.
Совокупность аксонов третьих нейронов составляет латеральную петлю. Следует отметить, что часть волокон поступает в латеральную петлю непосредственно от нейронов переднего и заднего улитковых ядер, транзитом проходя через трапециевидные ядра. Эти волокна прерываются на клетках, рассеянных по ходу латеральной петли. Данные клетки объединяются под названием "ядра петли".
Волокна латеральной петли поднимаются вверх. В области перешейка ромбовидного мозга они лежат поверхностно, располагаясь в проекции треугольника петли. Меньшая часть волокон латеральной петли заканчивается на клетках нижних холмиков среднего мозга. Большая часть волокон достигает второго подкоркового центра – ядер медиальных коленчатых тел промежуточного мозга. Третьим подкорковым центром являются срединные ядра таламуса, которые связаны с подкорковым чувствительным центром экстрапирамидной системы. Ядра подкорковых центров – это четвертые по счету нейроны слухового пути.
Аксоны клеток ядра нижнего холмика направляются в интеграционный центр среднего мозга, расположенный в верхнем холмике. От клеток последнего начинаются крышеспинномозговой и крыше-ядерный пути, которые направляются к двигательным ядрам передних рогов спинного мозга и двигательным ядрам черепных нервов. Названные пути проводят эфферентные нервные импульсы, осуществляющие безусловнорефлекторные двигательные реакции мускулатуры туловища, конечностей, головы и глазного яблока па внезапные слуховые раздражения.
Небольшое количество волокон латеральной петли направляется к нейронам срединных ядер таламуса. Аксоны клеток этих ядер передают информацию в интеграционный центр промежуточного мозга – медиальные ядра таламуса. Последние имеют связи с двигательными центрами экстрапирамидной, лимбической систем и гипоталамуса. Указанные структуры регулируют тонус мускулатуры, осуществляют разнообразные эмоциональные реакции в ответ на звуковые раздражения.
Аксоны клеток медиальных коленчатых тел проходят через заднюю ножку внутренней капсулы и затем, веерообразно рассыпаясь, направляются в среднюю часть верхней височной извилины – проекционный центр слуха. Путь от медиального коленчатого тела до верхней височной извилины носит название слуховой лучистости (коленчато-височный путь). В связи с тем что лишь небольшая часть волокон слухового пути от улитковых ядер идет по своей стороне, в проекционный центр слуха информация поступает преимущественно с противоположной стороны.
Кора полушарий большого мозга по принципу обратной связи оказывает влияние на подкорковые центры слуха и опосредованно – на кортиев орган. К последнему направляются аксоны от клеток заднего ядра трапециевидного тела (верхнего оливного ядра) – оливо-улитковый путь. Волокна этого пути заканчиваются на волосковых клетках. Корковые связи с кортиевым органом обеспечивают более точное восприятие звуков, восприятие шепотной речи и предохраняют от чрезмерно сильных звуков.
При поражении кортиева органа, улиткового нерва, переднего и заднего улитковых ядер возникает односторонняя глухота. Одностороннее поражение латеральной петли, медиального коленчатого тела и проекционного коркового центра сопровождается снижением слуха на оба уха. При локализации очага в ассоциативном центре слуха (задняя часть верхней височной извилины) могут наблюдаться слуховые галлюцинации.
2. Вестибулярный, или статокинетический, анализатор обеспечивает восприятие вестибулярных раздражений, проведение нервных импульсов до вестибулярных нервных центров, анализ и интеграцию поступившей в них информации. Функция вестибулярного анализатора заключается в получении информации о положении головы, ее движениях, а следовательно, и об изменениях положения тела в пространстве.
Рецепторы статокинетического анализатора располагаются в органе равновесия, которым являются три полукружных протока, эллиптический и сферический мешочки преддверия лабиринта внутреннего уха. В ампулах полукружных протоков рецепторы представлены клетками ампулярных гребешков, в мешочке и маточке – клетками пятнышек или отолитовых аппаратов. По своему строению рецепторные клетки являются первичночувствующими волосковыми сенсорными клетками.
Волосковые сенсорные клетки ампулярных гребешков полукружных протоков реагируют на движения головы или совместные движения головы и тела в определенной плоскости – горизонтальной, сагиттальной и фронтальной. Они воспринимают угловые ускорения, связанные с изменениями равновесия. Восприятие раздражения обусловлено инерционным смещением эндолимфы в полукружном протоке, находящемся в плоскости, соответствующей движению тела или головы. Волосковые сенсорные клетки эллиптического и сферического мешочков сигнализируют об изменениях положения тела по отношению к центру земной тяжести, воспринимают вертикальные ускорения, связанные с изменениями гравитационного поля.
У основания волосковых сенсорных клеток органа равновесия разветвляются периферические отростки биполярных клеток, тела которых располагаются в преддверном узле (первый нейрон). Последний находится на дне внутреннего слухового прохода височной кости.
Центральные отростки клеток преддверного узла образуют преддверный корешок преддверно-улиткового нерва. Он в области мосто-мозжечкового угла внедряется в вещество моста. В мосту волокна преддверного корешка направляются к вестибулярным ядрам (второй нейрон), расположенным в латеральном углу ромбовидной ямки.
Аксоны клеток вестибулярных ядер формируют несколько пучков, которые направляются в спинной мозг, в мозжечок, в состав медиального и заднего продольных пучков, а также в таламус. В спинной мозг идут часть аксонов клеток ядра Дейтерса и аксоны клеток ядра Роллера, образуя преддверно- спинномозговой путь (см. рис. 3.9). Этот путь в спинном мозге располагается на границе бокового и переднего канатиков и посегментно заканчивается на клетках двигательных ядер передних рогов спинного мозга. Он обеспечивает безусловнорефлекторное поддержание равновесия тела при вестибулярных нагрузках.
Часть аксонов клеток ядер Дейтерса и Швальбе, а также аксоны клеток ядра Бехтерева формируют преддверно-мозжечковый путь. Он проходит через нижние мозжечковые ножки и заканчивается на клетках коры червя мозжечка. Следует отметить, что клетки ядра Дейтерса имеют обратную связь с мозжечком в виде мозжечково-преддверного тракта. Через этот путь мозжечок оказывает опосредованное влияние на спинной мозг по преддверно-спинномозговому тракту.
Часть аксонов клеток ядра Дейтерса направляется в состав медиального продольного пучка своей и противоположной сторон и заканчивается на клетках интерстициального ядра (ядра Кахаля) и ядра задней спайки (ядра Даркшевича). Эти ядра ретикулярной формации обеспечивают связь органа равновесия с ядрами черепных нервов (III, IV, VI и XI пар), иннервирующими мышцы глазного яблока и мышцы шеи, обеспечивая сочетанный поворот головы и глаз.
Часть аксонов клеток ядра Дейтерса вступает в состав заднего продольного пучка и заканчивается на клетках заднего гипоталамического ядра. Последнее обеспечивает связь органа равновесия (через ядро Дейтерса) с вегетативными ядрами (III, VII, IX, X пар черепных нервов). Указанные связи позволяют объяснить появление вегетативных реакций (тошнота, рвота, побледнение кожи, похолодание конечностей, усиление потоотделения, усиление перистальтики органов желудочно-кишечного тракта, урежение пульса, снижение артериального давления, сужение зрачков и т.д.) в ответ на чрезмерные раздражения вестибулярного аппарата.
Меньшая часть аксонов клеток срединных ядер таламуса заканчивается на медиальных ядрах таламуса (подкорковый чувствительный центр экстрапирамидной системы), который осуществляет безусловно-рефлекторную регуляцию тонуса мускулатуры при вестибулярных нагрузках.
Сознательная оценка вестибулярных раздражений осуществляется только в коре полушарий большого мозга. Нервные импульсы к корковому концу анализатора поступают следующим образом. Часть аксонов клеток ядер Дейтерса и Швальбе переходит на противоположную сторону и формирует преддверно-таламический тракт. Этот тракт проходит в составе бульбарно-таламического тракта и заканчивается на клетках срединных ядер таламуса (третий нейрон). Аксоны клеток срединных ядер таламуса большей частью направляются через заднюю ножку внутренней капсулы в корковую часть вестибулярного анализатора (кора полушарий в области средней и нижней височных извилин).
Можно полагать, что в корковый центр вестибулярных функций информация поступает и опосредованно из центра двигательных функций, центра общей чувствительности и центра схемы тела. Наличие таких связей позволяет объяснить генерализацию эффекта в ответ на вестибулярные раздражения.
Заболевания органа равновесия (лабиринта), поражение вестибулярных ядер моста сопровождаются такими симптомами, как головокружение, расстройства равновесия и координации движений, нистагм (ритмичное подергивание глазных яблок), вегетативные расстройства. Поражение корковой части вестибулярного анализатора проявляется, кроме названных симптомов, нарушениями ориентации в пространстве. И, наконец, следует отметить, что в результате тренировок наступает привыкание к вестибулярным раздражениям.
3. Зрительный анализатор обеспечивает восприятие зрительных раздражений, проведение нервных импульсов до зрительных нервных центров, анализ и интеграцию поступившей в них информации.
Зрительный анализатор является одним из наиболее важных в познании внешнего мира. Через него поступает в кору полушарий большого мозга до 90% информации о предметах окружающей среды – их размерах, форме, цвете, пространственных отношениях, удаленности, направлении движения и т.д. Кроме того, зрительному анализатору также присуще свойство накопления, сохранения и узнавания ранее известной зрительной информации (зрительная память).
Рецепторы зрительного анализатора располагаются в зрительной части сетчатой оболочки глазного яблока. Сетчатка включает 10 слоев клеточных элементов, различных по своему строению и функциональному назначению. Рецепторами органа зрения являются палочки и колбочки, которые находятся в девятом по глубине залегания слое. Палочек насчитывается от 100 до 150 млн. Они обеспечивают видение предметов в сумерках (скотопическое зрение). Колбочек имеется только 5–7 млн. Они сосредоточены преимущественно в области желтого пятна сетчатой оболочки и отвечают за дневное зрение – воспринимают цвета (фототопическое зрение). Палочки и колбочки раздражаются под воздействием света – за счет ферментов (родопсина и йодопсина) в рецепторах протекают фотохимические реакции, преобразующие энергию светового раздражения в нервные импульсы. Нервные импульсы передаются на периферические отростки биполярных клеток, которые являются первыми нейронами зрительного пути и располагаются в сетчатке. Центральные отростки биполярных клеток имеют небольшую длину и синаптически заканчиваются на мультиполярных нервных клетках ганглиозного слоя сетчатой оболочки. Ганглиозные нейроны (вторые нейроны зрительного пути) – крупные и обычно контактируют одновременно с несколькими биполярными клетками. Аксоны ганглиозных клеток выходят на поверхность сетчатой оболочки, сближаются друг с другом в области диска зрительного нерва (слепое пятно) и формируют его ствол.
Зрительный нерв выходит из глазницы через зрительный канал и па основании мозга образует с одноименным нервом противоположной стороны зрительный перекрест – перекрещиваются только 2/3 нервных волокон, расположенных медиально. Эти волокна идут от внутренних отделов сетчатки, которая благодаря перекресту пучков света в хрусталике воспринимает зрительную информацию с латеральных сторон. Неперекрещивающиеся волокна, примерно 1/3, направляются в зрительный тракт своей стороны. Они идут от латеральных отделов сетчатки, которая воспринимает свет с носовой половины ноля зрения. Неполный перекрест зрительных путей имеет важное физиологическое значение. Он позволяет передавать нервные импульсы из каждого глаза в оба полушария. За счет этого обеспечивается бинокулярное стереоскопическое зрение и возможность синхронного движения глазных яблок.
Зрительный перекрест продолжается в зрительные тракты, содержащие волокна от одноименных половин сетчатки обоих глаз. Так, в составе правого зрительного тракта проходят неперекрещенные волокна от правой половины правого глаза и перекрещенные волокна от правой половины левого глаза. Следовательно, правый зрительный тракт проводит нервные импульсы, поступающие из носовой (медиальной) части поля зрения правого и латеральной части поля зрения левого глаза.
У латерального края ножки мозга зрительный тракт делится на три пучка, направляющихся к подкорковым центрам зрения – в верхние холмики крыши среднего мозга, в латеральные коленчатые тела и в задние ядра таламуса (рис. 4.6).
В верхние холмики идет небольшая часть волокон, обеспечивающих осуществление зрачкового рефлекса и безусловнорефлекторные двигательные реакции глазного яблока в ответ на световые раздражения. Аксоны клеток ядра верхнего холмика направляются к добавочным ядрам глазодвигательного нерва (ядра Якубовича), к клеткам ретикулярной формации, к двигательным ядрам III, IV и VI пар черепных нервов и к ядру Кахаля, от которого начинается медиальный продольный пучок. Клетки добавочных ядер посылают аксоны к ресничному узлу, который осуществляет парасимпатическую иннервацию мышцы, суживающей зрачок, и ресничной мышцы. Ресничная мышца обеспечивает аккомодацию глаза – способность ясно видеть предметы как на близком, так и на далеком расстоянии. Аккомодация осуществляется за счет изменения кривизны хрусталика. Мышца, расширяющая зрачок, получает симпатическую иннервацию. Благодаря наличию связей клеток верхнего холмика с парасимпатическим и симпатическим нервными центрами, иннервирующими мышцы радужки, регулируется поток света, поступающий на сетчатую оболочку глаза. При повышении освещенности зрачок суживается, и, наоборот, в темноте зрачок расширяется. Реакция зрачка на свет носит название "зрачковый рефлекс".
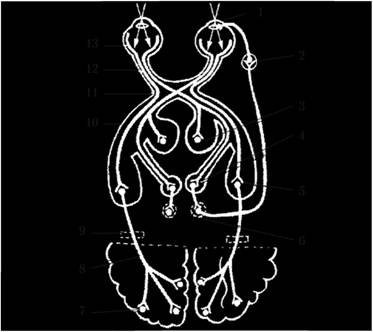
Рис. 4.6. Проводящий путь зрительного анализатора:
1 – ресничная мышца и мышца, суживающая зрачок; 2 – ресничный узел; 3 – задние ядра таламуса; 4 – верхний холмик среднего мозга; 5 – латеральное коленчатое тело; 6 – добавочные ядра глазодвигательного нерва; 7 – затылочная доля; 8 – зрительная лучистость; 9 – внутренняя капсула; 10 – зрительный тракт; 11 – зрительный перекрест; 12 – зрительный нерв; 13 – сетчатка
Связи нейронов ядра верхнего холмика с клетками двигательных ядер III, IV, VI пар черепных нервов обеспечивают двигательную реакцию мышц глазного яблока на световые раздражения. За счет этого осуществляется бинокулярное зрение – восприятие изображения одновременно обоими глазами.
Связи нейронов верхнего холмика с нейронами ядра Кахаля позволяют осуществлять согласованные движения глазных яблок и головы, что необходимо для поддержания равновесия тела.
Часть аксонов клеток ядра верхнего холмика направляется в интеграционный центр среднего мозга, который располагается также в верхнем холмике. От клеток последнего начинаются крыше-спинномозговой и крыше-ядерный пути. Эти пути проводят эфферентные нервные импульсы, осуществляющие безусловнорефлекторные двигательные реакции мускулатуры туловища, конечностей, головы и глазных яблок на внезапные световые раздражения.
Небольшое количество волокон зрительного тракта направляется к нейронам задних ядер таламуса. Аксоны клеток этих ядер передают зрительную информацию в интеграционный центр промежуточного мозга – медиальные ядра таламуса. Последние имеют связи с двигательными центрами экстра- пирамидной системы, лимбической системы и гипоталамуса. Указанные структуры регулируют тонус мускулатуры, осуществляют разнообразные эмоциональные реакции, изменяют работу внутренних органов в ответ на зрительные раздражения.
Главным подкорковым центром зрения является латеральное коленчатое тело (третий нейрон) – здесь заканчивается бо́льшая часть волокон зрительного тракта. Аксоны этих нейронов в виде компактного пучка проходят через заднюю ножку внутренней капсулы, затем веерообразно рассыпаются, образуя зрительную лучистость, или коленчато-шпорный путь. Он заканчивается на нейронах проекционного центра зрения (четвертый нейрон). Последний находится на медиальной поверхности затылочной доли по сторонам от шпорной борозды (поле 17). В глубине шпорной борозды заканчиваются волокна, проводящие импульсы от желтого пятна сетчатки (зона локализации колбочек).
Установлена проекция сетчатой оболочки на корковую часть анализатора зрения. Верхняя половина сетчатки проецируется на клетки верхнего края шпорной борозды, нижняя половина – на клетки нижнего края шпорной борозды. При этом нейроны в корковом конце анализатора группируются в колонки, расположенные радиально по отношению к поверхности коры. Каждая колонка осуществляет анализ и синтез только конкретного параметра зрительной информации – размера, формы, цвета, удаленности предмета и т.д. В прилежащих к проекционному центру зрения участках коры полушарий, в частности в пределах полей 18 и 19, осуществляются анализ, синтез и интеграция более сложной зрительной информации – узнавание ранее увиденного, элементы зрительного абстрагирования.
Нарушения функции зрительного анализатора весьма разнообразны и зависят от уровня поражения. При поражениях зрительного нерва, сопровождающихся вовлечением в патологический процесс всех его волокон, возникает полная слепота (амавроз) на один глаз. Частичные повреждения волокон или разрушение рефлексогенных зон сетчатой оболочки (отслойка, кровоизлияния) приводят к выпадению отдельных полей зрения (скотома).
Травматическое разрушение зрительного перекреста сопровождается полной слепотой на оба глаза. При опухолях гипофиза возможно сдавление внутренней части зрительного перекреста, которое сопровождается выпадением латеральных частей полей зрения (битемпоральная гемианопсия). Биназальная гемианопсия возникает при сдавлении зрительного перекреста с латеральных сторон.
Повреждение зрительного тракта, латерального коленчатого тела, зрительной лучистости или проекционного центра зрения приводит к гемианопсии (выпадение полей зрения с одноименных сторон на обоих глазах). Очаговое поражение в ассоциативных центрах зрения (поля 18 и 19) сопровождается неспособностью узнавания предметов, искаженностью их форм или зрительными галлюцинациями.
4. Вкусовой анализатор обеспечивает восприятие вкусовых раздражений, проведение нервных импульсов до вкусовых нервных центров, анализ и интеграцию поступившей в них информации.
Вкусовой анализатор играет важную роль в деятельности пищеварительной системы. Он представляет информацию о химическом составе, вкусе и качестве пищи. Кроме того, располагаясь в начальном отделе пищеварительной системы, вкусовой анализатор рефлекторно воздействует на железы (слюнные железы, железы желудочно-кишечного тракта, печень, поджелудочную железу), оказывая па них регулирующее влияние.
Вкусовые рецепторы находятся в полости рта и представлены вкусовыми клетками, которые входят в состав вкусовых почек (луковиц). У человека количество вкусовых почек колеблется от 3000 до 9000. Они располагаются большей частью на языке в области грибовидных, желобоватых и листовидных сосочков. Меньшее количество вкусовых почек находится в эпителии слизистой оболочки губ, мягкого неба, небных дужек, глотки, надгортанника. Совокупность вкусовых почек в полости рта составляет орган вкуса.
Вкусовая почка в центре имеет ямку, в которую попадают растворенные в слюне вещества. Вкусовые клетки функционально специализированы: сладкое воспринимается кончиком языка, кислое – его боковой поверхностью, горькое – корнем, соленое – всей поверхностью. Во вкусовых клетках химическое раздражение трансформируется в нервный импульс. Нервные импульсы синаптическим способом передаются на рецепторные окончания чувствительных нейронов.
Первые (чувствительные) нейроны вкусового пути представлены псевдоуниполярными клетками, расположенными в трех различных узлах – в коленчатом узле лицевого, в верхнем узле языкоглоточного и нижнем узле блуждающего нервов (рис. 4.7).
Периферические отростки псевдоуниполярных клеток коленчатого узла направляются к вкусовым почкам передних 2/3 языка в составе барабанной струны лицевого нерва. Центральные отростки в составе лицевого нерва входят в вещество моста в области мосто-мозжечкового угла.
Периферические отростки рецепторных нейронов верхнего узла языкоглоточного нерва направляются к вкусовым почкам задней трети языка в составе язычной ветви этого нерва. Центральные отростки формируют корешки языкоглоточного нерва и входят в вещество продолговатого мозга в области его задней латеральной борозды. Периферические отростки рецепторных нейронов нижнего узла блуждающего нерва направляются к вкусовым рецепторам глотки, надгортанника, неба в составе глоточных ветвей этого нерва, центральные отростки, как и у языкоглоточного нерва, вступают в вещество продолговатого мозга в области задней латеральной борозды.
Войдя в мозг, центральные отростки рецепторных клеток от всех трех узлов направляются к ядрам одиночного пути (второй нейрон). Аксоны клеток ядер одиночного пути в большинстве своем совершают перекрест и направляются в составе ядерно-таламического тракта к передним базальным ядрам таламуса (третий нейрон). Их аксоны большей частью проходят в составе таламо-коркового тракта к проекционному центру вкуса, расположенному в коре крючка и парагиппокампальной извилины (четвертый нейрон). Меньшая часть аксонов указанных ядер таламуса направляется к его медиальным ядрам. С последних информация поступает на подкорковые двигательные центры экстрапи- рамидной системы и к структурам лимбической системы, в частности в миндалевидное тело.
При поражении барабанной струны, лицевого нерва, язычной ветви языкоглоточного нерва возникают расстройства вкусовой чувствительности на одноименной стороне языка.
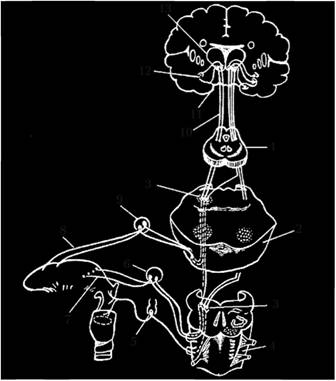
Рис. 4.7. Вкусовой путь:
1 – средний мозг; 2 – мост; 3 – ядра одиночного пути; 4 – продолговатый мозг; 5 – нижний узел блуждающего нерва; 6 – верхний узел языкоглоточного нерва; 7 – язычные ветви; 8 – барабанная струна; 9 – коленчатый узел; 10 – ядерно-таламический путь; 11 – крючок; 12 – гиппокамп; 13 – таламус
При поражении передних базальных ядер таламуса, таламо-коркового пучка, проекционного центра вкуса отмечается частичное снижение вкуса на противоположной стороне.
При поражении миндалевидного тела больной не может обозначить вкус, хотя его ощущает (вкусовая агнозия). Полная утрата вкуса носит название "агевзия", снижение вкуса – "гипогевзия", извращение вкусовых ощущений – "дисгевзия".
5. Обонятельный анализатор обеспечивает восприятие обонятельных раздражений, проведение нервных импульсов до обонятельных нервных центров, анализ и интеграцию поступившей в них информации.
Обонятельный анализатор в жизни человека играет важную роль. Он позволяет контролировать качество вдыхаемого воздуха, качество принимаемой пищи, в совокупности с другими анализаторами позволяет ориентироваться в окружающей среде. Большое значение имеет наличие у людей обонятельной памяти, обеспечивающей возможность узнавать ранее встречавшиеся запахи.
Рецепторы, воспринимающие обонятельные раздражения, располагаются в обонятельной области слизистой оболочки полости иоса. Они занимают площадь до 10 см2 в пределах верхнего носового хода, верхней носовой раковины и верхней части перегородки полости иоса. Рецепторы представлены разветвлениями периферических отростков биполярных клеток слизистой оболочки. Количество биполярных обонятельных нейронов (рецепторных) у человека составляет до 10 млн. Периферические отростки у этих клеток короткие и заканчиваются булавовидными утолщениями, выступающими над поверхностью слизистой оболочки.
На каждом булавовидном утолщении находятся 10–15 обонятельных волосков, которые погружены в слой слизи. Пахучие вещества, попадая с потоком воздуха в верхний носовой ход, растворяются в слизи. Обонятельные волоски взаимодействуют с молекулами пахучих веществ и трансформируют энергию химического раздражения в нервные импульсы.
Центральные отростки рецепторных клеток собираются в пучки по 15–20 волокон, формируя обонятельные нервы, которые через отверстия в решетчатой пластинке решетчатой кости проникают в полость черепа. Обонятельные нервы идут к обонятельной луковице, где вступают в синаптические контакты с дендритами митральных клеток (второй нейрон).
Аксоны митральных клеток объединяются в составе обонятельного тракта и вблизи обонятельного треугольника распадаются на три пучка – медиальный, промежуточный и латеральный (рис. 4.8). Волокна медиального пучка направляются через переднюю спайку в обонятельный тракт противоположной стороны и заканчиваются на митральных клетках обонятельной луковицы.
Волокна промежуточного пучка образуют синаптические окончания на нейронах обонятельного треугольника, переднего продырявленного вещества и нейронах ядра прозрачной перегородки. Часть волокон промежуточного пучка через переднюю спайку мозга следует к названным структурам противоположной стороны.
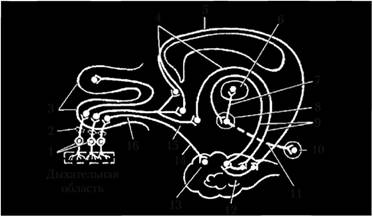
Рис. 4.8. Проводящий путь обонятельного анализатора:
1 – биполярные клетки; 2 – продырявленная пластинка; 3 – обонятельная луковица; 4 – медиальный обонятельнокорковый тракт; 5 – пояс; 6 – передние ядра таламуса; 7 – сосцевидно-таламический пучок; 8 – сосцевидное тело; 9 – свод; 10 – верхний холмик среднего мозга; 11 – сосцевидно-покрышечный пучок; 12 – парагиппокампальная извилина; 13 – крючок; 14 – латеральный обонятельно-корковый тракт; 15 – обонятельный треугольник; 16 – обонятельный тракт
Волокна наиболее крупного латерального пучка направляются непосредственно к корковому концу обонятельного анализатора и заканчиваются на нейронах старой коры полушарий большого мозга в крючке и парагиппокампальной извилине, образуя латеральный обонятельно-корковый тракт. Таким образом, особенностью обонятельного пути является то, что нервные импульсы первоначально поступают не в подкорковые центры обоняния, а в кору полушарий большого мозга – крючок и парагиппокампальную извилину.
Аксоны третьих нейронов, расположенных в обонятельном треугольнике и переднем продырявленном веществе, также направляются в корковый конец обонятельного анализатора – крючок и парагиппокампальную извилину, образуя медиальный обонятельно-корковый тракт. В эти участки коры полушарий височной доли они попадают различными путями: в составе медиального тракта, расположенного на дорсальной поверхности мозолистого тела; в составе свода и поясного пучка сводчатой извилины.
В подкорковые центры обоняния – сосцевидные тела – нервные импульсы поступают из проекционного центра обоняния (крючок и парагиппокампальная извилина). Названные структуры связывают между собой проекционные волокна, проходящие в составе свода. Следует отметить, что имеются два подкорковых центра обоняния. Кроме сосцевидных тел, подкорковым центром обоняния также являются передние ядра таламуса. Между собой подкорковые центры связаны сосцевидно-таламическим пучком (пучком Вик д'Азира).
Аксоны клеток передних ядер таламуса, в свою очередь, формируют два пучка. Один пучок через заднюю ножку внутренней капсулы направляется к проекционному обонятельному центру новой коры полушарий большого мозга, расположенному на вентральной поверхности лобных долей. Меньшая часть волокон идет в интеграционный центр промежуточного мозга – в медиальные ядра таламуса. Указанные центры связаны с двигательными экстрапирамидными центрами, лимбическими структурами и ядрами ретикулярной формации.
Медиальные ядра сосцевидных тел связаны с интеграционным центром среднего мозга – верхними холмиками. От нейронов верхнего холмика начинаются крыше-спинномозговой и крыше-ядерный пути. Эти пути проводят афферентные нервные импульсы, осуществляющие безусловно-рефлекторные двигательные реакции мускулатуры туловища, конечностей, головы и глазных яблок на внезапные сильные запахи.
При заболеваниях слизистой оболочки полости носа, опухолях основания мозга и лобной доли отмечается снижение обоняния (гипосмия) или полная его потеря (аносмия). При аллергических состояниях с вовлечением слизистой оболочки полости носа нередко отмечается обострение обоняния (гиперосмия). Поражение височной доли в области крючка и парагиппокампальной извилины сопровождается обонятельными галлюцинациями.