Методы определения генотипического состава популяций фитопатогенов
Фитопатогенные организмы обладают двумя важнейшими признаками — вирулентностью и агрессивностью.
Вирулентность — это качественный признак, т.е. способность определенного генотипа паразита заражать определенный генотип растения- хозяина. Вирулентностью определяется специализация паразитов в рамках семейств растений-хозяев (злаки не болеют фитофторозом картофеля, а пасленовые — спорыньей), родственных видов (специализированные формы возбудителя стеблевой ржавчины Puccinia graminis могут быть патогенными для пшеницы, овса, ячменя, мятлика и других родов злаков) и даже отдельных сортов (раса 1 возбудителя фитофтороза картофеля патогенна для сорта картофеля "Агрономический", но не для сорта "Домодедовский", а раса 3 — наоборот, вирулентна для сорта "Домодедовский", но не для сорта "Агрономический") (см. рис. 2.6).
Агрессивность — количественный признак, т.е. степень патогенности вирулентного штамма. Между вирулентностью и агрессивностью существуют определенные связи: если паразит не имеет вирулентности к какому-либо виду или сорту растения, у него нет и аїрессивности к этому растению, но если вирулентность есть, то агрессивность разных штаммов вирулентного паразита может быть высокой или низкой, т.е. они способны вызывать заболевание в сильной или слабой степени. Таким образом, вирулентность паразита есть зеркальное отображение вертикальной устойчивости растения, а агрессивность — горизонтальной. Сходство наблюдается и в наследовании этих признаков: вирулентность контролируется олигогенами, а агрессивность — полигенами.
Методы анализа вирулентности паразитов
Изменение вирулентности часто приводит к тяжелым последствиям — массовым заболеваниям многих ранее устойчивых сортов. Поэтому анализ генов вирулентности в популяциях паразитов, структуры и динамики вирулентных рас имеет очень большое практическое значение. Методы определения вирулентных рас ржавчинных грибов злаков впервые были разработаны в начале XX в. Стекменом. Он предложил набор из 12 сортов пшеницы, по реакции на заражение которых следует определять расы Puccinia graminis. Два сорта этого набора, расположенные по краям списка ("Литтл Клаб" и "Капли"), не являются дифференциаторами, так как первый универсально восприимчив, поражается всеми американскими изолятами возбудителя стеблевой ржавчины, а второй — универсально устойчив и не поражается ни одним изолятом. Они используются в наборе в качестве контролей. Остальные 10 сортов поражаются одними изолятами и не поражаются другими, т.е. способны дифференцировать популяцию. По методике Стекмена листья с пустулами ржавчины собирают в поле; в лаборатории спорами из отдельных пустул (мононустульные штаммы) заражают листья всходов сорта "Литтл Клаб" (для размножения каждого штамма, так как этот сорт поражают все изоляты) и полученным материалом заражают листья всходов остальных сортов-дифференциаторов. После образования симптомов болезни в журнале записывают реакцию растений на заражение [1]. Первый штамм ржавчины, проанализированный подобным образом, получил название расы 1. Второй, который отличался от первого реакцией на каком-либо дифференциаторе, был назван раса 2 и т.д. Система Стекмена получила широкое распространение. Подобным образом, только с использованием иных сортов-дифференциаторов, стали определять расы других грибов. Однако вскоре выяснилось, что эта система имеет существенные недостатки:
1) номенклатурные трудности. Название рас по Стекмену — это лишь порядковый номер идентификации, никоим образом не связанный со свойствами расы. Сейчас в мире описаны несколько сотен рас возбудителя стеблевой ржавчины, и чем отличается раса 256 от расы 312, не может сказать ни один исследователь; надо обращаться к справочным таблицам;
2) метод определения рас на стандартном наборе не отражает процесс эволюции рас в природе. Стекмен выбрал для дифференциации сорта пшеницы, используемые американскими селекционерами в период его работы (20-е гг. XX в.). В европейской селекции эти сорта никогда широко не использовались, да и в США интересы селекционеров со временем менялись, в то время как дифференциаторы оставались неизменными.
Например, в 1940-х гг. в ряде штатов был сильно поражен ржавчиной ранее устойчивый сорт "Ли". Фитопатологи, собрав пустулы с растений этого сорта, определили, что он поразился расой 15, которая была давно известна и к которой этот сорт был устойчив. Значит, это — новая раса, которая отличается от известных рас по реакции сорта "Ли" (не входящего в список дифференциаторов), но не отличается но реакции сортов стандартного набора. Назвали ее биотип 15В, и для отличия его от старого биотипа 15А пришлось к набору дифференциаторов прибавить сорт "Ли". Прошло несколько лет, и поразился сорт "Селкирк", ранее устойчивый к стеблевой ржавчине. Анализ показал, что его заразил биотип 15В, к которому ранее "Селкирк" был устойчив. Новую расу назвали 15В-2 (в отличие от старого биотипа 15В-1), и для его дифференциации пришлось к набору Стекмена добавить еще один сорт — "Селкирк". И этому процессу прибавлений нет конца. При этом каждый добавленный дифференциатор увеличивает емкость шкалы (максимальное число рас, которое можно определить с помощью данной шкалы) вдвое — вместо 2n — 2n+1. В Советском Союзе на дополнительном наборе из пяти сортов пшеницы, прибавленном к стандартному набору дифференциации рас бурой ржавчины, в лаборатории украинского фитопатолога М. П. Лесового было обнаружено 46 биотипов 77-й расы [2].
Ситуация с определением вирулентности существенно поменялась после работ шотландского фитопатолога и селекционера У. Блэка, который совместно с коллегами из Голландии и США разработан метод оценки рас возбудителя фитофтороза картофеля. Шкала Блэка содержит любой восприимчивый к фитофторе сорт культурного картофеля Solanum tuberosum и набор гибридов с диким мексиканским картофелем Solanum demissum, получившим от дикаря разные известные гены вертикальной устойчивости — R1, R2, R3, R4 и их сочетания. Число комбинаций из четырех генов составляет 24 = 16. Обозначения рас, заражающих те или иные дифференциаторы, показано в табл. 3.3.
Таблица 33
Генетическая дифференциация рас Phytophthora infestans
|
Раса |
Ген устойчивости картофеля |
|||||||||||||||
|
г |
R1 |
R2 |
R3 |
R4 |
R1 R2 |
R1 R3 |
R1 R4 |
R2 R3 |
R2 R4 |
R3 R4 |
R1 R2 R3 |
R1 R2 R4 |
R1 R3 R4 |
R2 R3 R4 |
R1 R2 R3 R4 |
|
|
0 |
+ |
|||||||||||||||
|
1 |
+ |
+ |
||||||||||||||
|
2 |
+ |
- |
+ |
|||||||||||||
|
3 |
+ |
+ |
||||||||||||||
|
4 |
+ |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
1.2 |
+ |
+ |
+ |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
1.3 |
+ |
+ |
- |
+ |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
- |
|
1.4 |
+ |
+ |
- |
- |
+ |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
- |
- |
|
2.3 |
+ |
- |
+ |
+ |
+ |
|||||||||||
|
2.4 |
+ |
- |
+ |
- |
+ |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
- |
|
3.4 |
+ |
- |
- |
+ |
+ |
- |
- |
- |
- |
- |
+ |
- |
- |
- |
- |
- |
|
1.2.3. |
+ |
+ |
+ |
+ |
- |
+ |
+ |
- |
+ |
- |
- |
+ |
- |
- |
- |
- |
|
1.2.4. |
+ |
+ |
+ |
- |
+ |
+ |
- |
+ |
- |
+ |
- |
- |
+ |
- |
- |
- |
|
1.3.4 |
+ |
+ |
- |
+ |
+ |
- |
+ |
+ |
- |
- |
+ |
- |
- |
+ |
- |
- |
|
2.3.4 |
+ |
- |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
- |
- |
+ |
- |
|
1.2.3.4 |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
Преимущества данной шкалы:
1) дифференциация, в соответствии с концепцией "ген против гена", опущена с генотипа (сорта) растения до отдельного гена ВУ. Это значит, что если раса 1 заражает дифференциатор, имеющий ген R1, то отсюда следует, что она будет заражать все сорта, имеющие только этот ген устойчивости, и при высокой концентрации данной расы в популяциях не следует использовать этот ген в селекции новых сортов, так как они будут поражаться;
2) для дифференциации не обязательно использовать все 16 дифференциаторов; достаточно иметь только пять (универсально восприимчивый сорт для контроля и четыре носителя каждого гена поодиночке). Если какой-либо штамм заражает дифференциаторы, имеющие гены R1 и R2, он априори будет заражать дифференциатор, имеющий оба эти гена. Таким образом, на пяти сортах можно определить 16 рас патогена;
3) номенклатура рас отражает их свойства. Каждая цифра в обозначении расы указывает на то, какой ген устойчивости не является эффективным, т.е. не защищает растение от данной расы. Конечно, такая номенклатура может быть слишком громоздкой: в период работ Блэка было известно четыре гена устойчивости к фитофторозу, и самая вирулентная раса имела обозначение из четырех цифр, сейчас описано 11 генов, поэтому обозначения высоко вирулентных рас фитофторы картофеля стали чересчур длинными. А у многих ржавчинных грибов, поражающих
злаки, известны десятки генов устойчивости, и номенклатура рас становится совсем неудобоваримой. Поэтому предложены разные двоичные и троичные системы записи рас, вполне пригодные для написания и расшифровки.
Конечно, и генетическая дифференциация не защищает от курьезов, подобных случившемуся с расой 15 стеблевой ржавчины пшеницы. Когда были обнаружены гены устойчивости к фитофторозу R5, R6 и т.д., то оказалось, что некоторые авирулентные расы, обозначаемые как 0, вирулентны для сортов с этими генами. Для их дифференциации надо, подобно сорту "Ли", добавлять в набор дифференциаторов сорта с новыми генами.
Недостаток любого способа дифференциации рас фитопатогенных грибов — низкая производительность работы. По разработанной Стекменом методике необходимо собрать в поле средний образец зараженных растений, материалом каждой пустулы заразить универсально восприимчивый сорт и размножить па нем инфекционный материал, заразить им сорта- дифференциаторы, растущие в горшках, и после появления симптомов болезни описать реакции на заражение. Следовательно, чтобы проанализировать расовый состав на одном ноле, необходимо затратить много времени и иметь большие площади теплиц. При недостатке рабочих рук и тепличных площадей за сезон в одной лаборатории обычно анализируют не больше 100—200 образцов.
Выборка из 100 пустул означает, что максимальная разрешающая способность анализа не превышает 1% (на самом деле гораздо ниже). Поскольку над одним гектаром пшеницы при средней зараженности ежедневно формируется 1012 спор ржавчины, 1% от 1012 составляет 10,(), т.е. 10 млрд спор. Поэтому "редкие" расы, отмеченные всего одним образцом, на самом деле вовсе не редкие, они встречаются в популяциях с частотой до 10 млрд на гектаре посевов. Действительно редкие расы при существующей методике анализа вообще не выявляются. А для изучения эволюции популяций, тенденций в их изменениях, необходимо иметь представления именно о редких генотипах и их динамике в пространстве и во времени. Поэтому все модификации определения рас, направленные па ускорение методов определения, очень важны. Например, для определения рас возбудителя фитофтороза картофеля проводят заражение не целых растений, а отдельных листьев, помещенных во влажные камеры (чашки Петри с увлажненной фильтровальной бумагой на дне). Расы ржавчинных грибов злаков таким методом определять нельзя, так как чтение реакций на заражение проводят не через три — пять дней после заражения, как при фитофторозе картофеля, а через 7—10 дней. За этот срок отрезки листьев желтеют, и реакция на заражение может меняться.
Канадские фитопатологи К. Персон и Д. Самборский предложили помещать отрезки листьев злаков на фильтровальную бумагу или вату, смоченную раствором фитогормона бензимидазола, который защищает хлоропласты от распада и тем самым более длительное время сохраняет ювенильное состояние отрезков листьев (рис. 3.5).
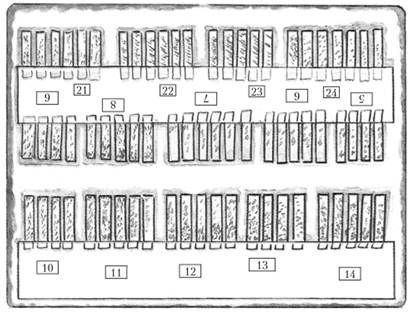
Рис. 3.5. Заражение изолятами возбудителя бурой ржавчины отрезков листьев растений-дифференциаторов на растворе бензимидазола [3]:
отрезки листьев пшеницы разложены в кювете со смоченной бензимидазолом фильтровальной бумагой на дне; каждый отрезок листа в кювете заменяет вазон с соответствующим сортом, используемый при стандартном методе определения рас
Замена целых растений в горшках на отрезки листьев позволяет на порядок увеличить число определяемых образцов. Однако и при этом методе остается необходимым размножение каждой монопустульной культуры на универсально восприимчивом сорте.
Метод анализа сборного образца спор, разработанный Л. А. Михайловой и К. В. Квитко, увеличивал разрешающую способность метода определения рас практически безгранично [4]. Они предложили смывать споры, образовавшиеся па зараженных листьях, в одну колбу (получать сборный образец спор), определять число спор в миллилитре в счетной камере Горяева и разбрызгивать дозированное количество споровой суспензии на чашки Петри, дно которых полностью покрыто отрезками листьев сорта-дифференциатора, помещенных на фильтровальную бумагу или вату, смоченную раствором бензимидазола (рис. 3.6).
Число пустул, образовавшихся на чашках с листьями универсально восприимчивого сорта, укажет на относительную концентрацию инфекционных единиц в инокулюмс. Сравнение числа пустул, образующихся па чашке с листьями сорта, имеющего определенный ген устойчивости, с числом пустул на чашке с универсально восприимчивым сортом укажет на процент вирулентных к этому сорту рас.

Рис. 3.6. "Газон" из отрезков листьев пшеницы на растворе бензимидазола, опрыснутый суспензией урединиоспор Puccinia triticina [5]:
па отрезках листьев видны пустулы ржавчины, число которых соответствует концентрации инфекционных единиц в инокулюме. Метод позволяет определить относительные частоты клонов паразита, вирулентных к разным сортам
Данный метод часто используют для анализа рас возбудителя бурой ржавчины пшеницы Puccinia triticina. Но его модификации можно использовать для определении концентрации вирулентных клонов и у возбудителей других болезней, вызывающих локальные пятна на листьях (гельминтоспориозы злаков, фитофтороз картофеля).