Кольчатые черви
Кольчатые черви, представляющие весьма обширную группу, являются эволюционными потомками плоских червей. Наиболее изученными из них являются живущие в морях многощетинковые черви – полихеты и малощетинковые черви – олигохеты. Наиболее известными представителями олигохет являются дождевой червь и пиявка. Характерным признаком строения кольчатых червей является внешняя и внутренняя метамерия: их тело состоит из нескольких, большей частью идентичных, сегментов, каждый из которых содержит комплект внутренних органов, в частности пару симметрично расположенных ганглиев с нервными комиссурами. В результате нервная система кольчатых червей имеет вид "нервной лестницы".
Особое место занимают представители класса малощетинковых – дождевые черви, на которых проводились основные опыты, связанные с изучением их реакций на разнообразные агенты среды и с выработкой условных рефлексов. Нервная система дождевых червей представлена в виде нервных узлов – ганглиев, расположенных вдоль всего тела в виде симметричной цепочки. Каждый узел состоит из грушевидных клеток и густого сплетения нервных волокон. От этих клеток к мышцам и к внутренним органам отходят двигательные нервные волокна. Под кожным покровом червя расположены чувствительные клетки, которые соединяются своими отростками – чувствительными волокнами – с нервными узлами. Нервная система подобного типа называется цепочечной, или ганглиозной. Тело дождевого червя состоит из ряда сегментов-члеников. Каждый сегмент имеет свой собственный нервный узел и может отвечать па раздражение, будучи совершенно отделен от всего остального тела, но все узлы соединены между собой перемычками, и организм действует как единое целое. Головной узел нервной системы, расположенный в верхней части головы, получает и перерабатывает наибольшее количество раздражений. Он устроен значительно сложнее, чем все остальные узлы нервной системы червя.
Движения кольчатых червей
Двигательная активность кольчатых червей отличается большим многообразием и достаточной сложностью. Обеспечивается это сильно развитой мускулатурой, состоящей из двух слоев: внешнего, состоящего из кольцевых волокон, и внутреннего – из мощных продольных мышц. Последние простираются, несмотря на сегментацию, от переднего до заднего конца туловища. Ритмичные сокращения продольной и кольцевой мускулатуры кожно-мышечного мешка обеспечивают движения. Червь ползет, вытягивая и сокращая, расширяя и сужая отдельные части своего тела. У дождевого червя вытягивается и сужается передняя часть тела, затем тоже самое происходит последовательно со следующими сегментами. В результате по телу червя пробегают "волны" сокращений и расслаблений мускулатуры.
У кольчатых червей впервые в эволюции животного мира появляются подлинные парные конечности: на каждом сегменте имеется по паре выростов, получивших название параподий. Они служат органами передвижения и снабжены специальными мышцами, двигающими их вперед или назад. Зачастую параподии имеют ветвистое строение. Каждая ветвь снабжена опорной щетинкой и, кроме того, венчиком из щетинок, имеющих у разных видов различную форму. От параподий отходят и щупальцевидные органы тактильной и химической чувствительности. Особенно длинными и многочисленными последние являются на головном конце, где на спинной стороне располагаются глаза (одна или две пары), а в ротовой полости или на особом выпячиваемом хоботке – челюсти. В захвате пищевых объектов могут участвовать и нитевидные щупальца на головном конце червя.
Поведение кольчатых червей
Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная их масса ведет придонный образ жизни в прибрежных зонах, где селится среди колоний других морских организмов или в расщелинах скал. Многие виды живут временно или постоянно в трубках, которые в первом случае периодически покидаются их обитателями, а затем вновь разыскиваются. Особенно хищные виды регулярно отправляются из этих убежищ на "охоту". Трубки строятся из песчинок и других мелких частиц, которые скрепляются выделениями особых желез, чем достигается большая прочность построек. Неподвижно сидящие в трубках животные ловят свою добычу (мелкие организмы), подгоняя к себе и процеживая воду с помощью венчика щупальцев, который высовывается из трубки, или же прогоняя сквозь нее поток воды (в этом случае трубка открыта на обоих концах).
В противоположность сидячим формам свободноживущие черви активно разыскивают свою пищу, передвигаясь по морскому дну: хищные виды нападают на других червей, моллюсков, ракообразных и иных сравнительно крупных животных, которых хватают челюстями и проглатывают; растительноядные отрывают челюстями куски водорослей; другие черви (их большинство) ползают и роются в придонном иле, проглатывают его вместе с органическими остатками или собирают с поверхности дна мелкие живые и мертвые организмы.
Малощетинковые черви ползают и роются в мягком грунте или придонном иле, некоторые виды способны плавать. Во влажных тропических лесах некоторые малощетинковые кольчецы вползают даже на деревья. Основная масса малощетинковых червей питается дейтритом, всасывая слизистый ил или прогрызаясь сквозь почву. Но существуют и виды, поедающие мелкие организмы с поверхности грунта, процеживающие воду или отгрызающие куски растений. Несколько видов ведут хищный образ жизни и захватывают мелких водных животных, резко открывая ротовое отверстие. В результате добыча всасывается с потоком воды.
Пиявки хорошо плавают, производя туловищем волнообразные движения, ползают, роют ходы в мягком грунте, некоторые передвигаются по суше. Помимо кровососущих существуют также пиявки, которые нападают на водных беспозвоночных и проглатывают их целиком. Наземные пиявки, обитающие во влажных тропических лесах, подстерегают свои жертвы на суше, в траве или на ветках деревьев и кустарников. Они могут довольно быстро двигаться. В передвижении наземных пиявок по субстрату большую роль играют присоски: животное вытягивает туловище, затем присасывается к субстрату головной присоской и притягивает к ней задний конец туловища, одновременно сокращая его, затем присасывается задней присоской и т.д.
Экспериментальное изучение поведения кольчатых червей
Дождевые или земляные черви широко распространены на территории всего земного шара. Эти животные играют огромную роль в почвообразовании, поэтому они издавна привлекают к себе пристальное внимание ученых самых разных профилей. Достаточно хорошо изучено и их поведение. Так, жизнедеятельность дождевых червей подробно описывал еще Ч. Дарвин. В ходе его опытов выяснилось, что они по-разному реагируют на зрительные, осязательные, обонятельные и температурные раздражители. Р. Йеркс и ряд других ученых исследовали у дождевых червей способность к образованию простейших навыков. Для этой цели чаще всего использовалась методика выработки оборонительных условных реакций в Т-образном лабиринте. Черви обучались поворачивать в правый или левый рукав лабиринта. Безусловным раздражителем служил переменный ток различной интенсивности, а условным – сам лабиринт, элементы которого, вероятно, воспринимались проприоцептивной и тактильной афферентациями. Критерием выработки рефлекса служило увеличение числа поворотов в рукав лабиринта, где животные не подвергались электрической стимуляции. В опытах Р. Йеркса черви обучались правильному выбору стороны после 80–100 сочетаний (рис. 15.3).
Наличие органов чувств помогает дождевым червям различать простейшие формы. Так, в процессе запасания пищи они схватывают двойные сосновые иглы за основание, а опавшие листья за верхушки, за которые они втягивают их в свою норку.
Еще более четкие условные рефлексы удается вырабатывать у многощетинковых червей – полихет. Так, у нереисов удавалось выработать устойчивые условные рефлексы на тактильное раздражение, пищу, свет и вибрацию Анализ результатов показал, что у полихет вырабатываются реакции, обладающие всеми основными свойствами истинных условных рефлексов: возрастание числа положительных ответов от опыта к опыту, высокий максимальный процент положительных реакций (до 80–100) и длительность их сохранения (до 6–15 дней).
Весьма существенно, что выработанная реакция угасала при отсутствии подкрепления и самопроизвольно восстанавливалась.
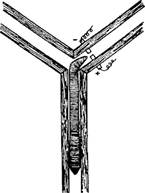
Рис. 15.3. Обучение червя в Т-образном лабиринте
Выявленные закономерности условно-рефлекторной деятельности полихет коррелируют с относительно дифференцированным мозгом животных. Таким образом, истинные условные рефлексы как один из достаточных совершенных механизмов, определяющих приобретенное поведение, впервые в эволюции, по-видимому, появляются у кольчатых червей[1].