Связь с клиникой: переливание крови и пересадка тканей
Совместимость поверхностных клеточных антигенов во многом определяет успех переливания крови или трансплантации тканей. Наиболее важные поверхностные антигены, которые определяют успех переливания крови, это антигены групп крови.Их антигенные свойства обусловлены тонкой структурой углеводных остатков гликопротеинов и гликолипидов на поверхности эритроцитов.
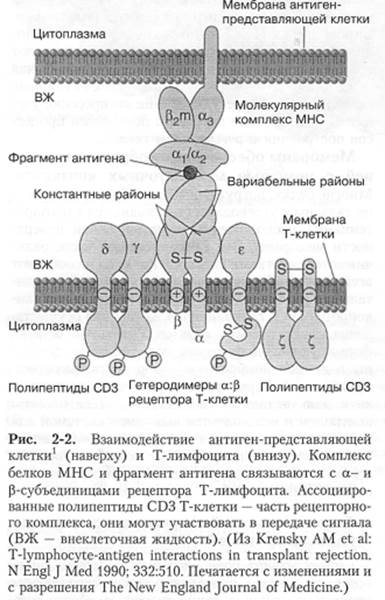
Второй комплекс антигенов, называемый трансплантационными антигенами или антигенами гистосовместимости,определяет индивидуальность поверхностей клеток. В отличие от антигенов групп крови (определяющих антигенность как жидкой среды организма, так и клеточной поверхности), антигены гистосовместимости размещены лишь на поверхности клеток. Антигенными детерминантами служат полипептидные цепи группы трансмембранных белков, кодируемых в геноме млекопитающих генами главного комплекса гистосовместимости (МНС).
Таким образом, посредством антигенов гистосовместимости и антигенов групп крови плазматическая мембрана клеток обеспечивает генетическую индивидуальность клетки. Этот комплекс неоценим для защиты организма. Особенно важно, что иммунный ответ Т-лимфоцита активируется при контакте с клеточным поверхностным антигеном (АГ), который представляется Т-лимфоциту молекулой МНС.
Строение и сборка мембран
Локализация синтеза
Синтез белка происходит в основном в двух местах: в цитозоле и на мембране эндоплазматического ретикулума (ЭР). Белки, которые высвобождаются в цитоплазму для дальнейшего пребывания во внутренней среде клетки, синтезируются на рибосомах,локализованных в цитозоле.Фактически все рибосомы располагаются в цитозоле; однако, до того как они соединятся с мРНК, рибосомы существуют в виде двух основных рибосомных субъединиц, рибонуклеопротеиновых комплексовс константами седиментации 40S и 60S.
После выхода мРНК в цитоплазму через поры ядерной мембраны образуется начальный комплекс, состоящий из рибосомной 40S субъединицы и мРНК. Как только данный комплекс сформируется, к нему присоединяется рибосомная субъединица 60S, в результате образуется функциональная рибосома, которая немедленно начинает транслировать на мРНК полипептид.
Если рибосома встречает нуклеотидную последовательность, кодирующую аминокислотную цепь, сигнализирующую, что транслируемый белок является мембранным или секретируемым, то трансляция прекращается до встраивания начального конца белка в эндоплазматический ретикулум (сигнальная гипотеза).
Результаты исследований синтеза белка (в том числе синтеза на полирибосомах, ассоциированных с ЭР, — «мембрано-связанных рибосомах»), показали, что все интегральные белкисинтезируются на мембране ЭР, включая белки плазматической мембраны. На рисунке показаны клеточные органеллы, участвующие в синтезе и процессинге белка.

Липиды мембрантакже синтезируются на ЭР и переносятся в плазматическую мембрану. Процесс переноса белков и липидов из ЭР к плазматической мембране осуществляется транспортными пузырьками.
Важным следствием синтеза компонентов плазматической мембраны на ЭР является тот факт, что эти компоненты транспортируются в форме готовых мембран, т.е. в мембранных пузырьках.Таким образом, важно помнить, что мембраны происходят только из предсуществующих мембран. Мембраны органелл клетки имеют сходное строение; однако по белково-липидному составу они значительно отличаются от плазматической мембраны и друг от друга.