Система выделительных тканей Выделительные системы растений (железистые волоски, выделительные ходы, млечники и т.д.)
Выделительные системы в жизнедеятельности растений играют важную, разнообразную роль. В процессе их жизнедеятельности образуется ряд веществ с разнообразной химической природой, не участвующих в дальнейшем метаболизме. Выделительные структуры растений характеризует, то что клетки выделительных тканей паренхимные, тонкостенные. Их ультраструктура связана с секретирующим веществом. В тканях, где синтезируются эфирные масла, смолы, каучук, имеется хорошо развитый агранулярный ЭР, слизи - аппарат Гольджи. Выделительные ткани подразделяются на ткани внешней (наружной) и внутренней секреции. Наружные выделительные структуры (внешняя секреция). Связаны эволюционно с покровными тканями. Они представлены следующими структурами: железистые волоски, желёзки, переваривающие желёзки, нектарники, осмофоры, гидатоды. Железистые волоски и желёзки представляют собой трихомы эпидермы. Они состоят из живых клеток, обычно имеют удлиненную ножку из одной или нескольких клеток и одно- или многоклеточную головку. Клетки головки выделяют секрет под кутикулу. При разрыве кутикулы вещество изливается наружу, после чего может образовываться новая кутикула и накопиться новая капля секрета. Железистые волоски цветков герани, листьев и цветков душистого табака выделяют эфирные масла; сидячие головчатые волоски, образующие мучнистый налёт на листьях мари и лебеды, - воду и соли. Желёзки отличаются от волосков короткой ножкой из несекретирующих клеток и многоклеточной головкой. Они характерны у мяты, лаванды, полыни, черной смородины. Внутренние выделительные структуры (внутренняя секреция). Вырабатывают и накапливают вещества, остающиеся внутри растения. Это могут быть отдельные секреторные клетки, рассеяные среди других тканей, как идиобласты. Они содержат различные вещества, особенно часто оксалат натрия в виде одиночных кристаллов, друз или рафид, бальзамы, танины, слизи и др. лечники – особый тип выделительной ткани. Это живые клетки (нечленистые млечники) или ряды слившихся клеток (членистые млечники), пронизывающие всё растение. В зрелом млечнике протопласт занимает постенное положение, полость млечника занята млечным соком – латексом, млечные клетки неодревесневающие, эластичные. Латекс представляет собой эмульсию белого, реже оранжевого или красного цвета. Жидкая основа латекса – клеточный сок, в котором растворены или взвешены углеводы (крахмальные зерна у молочайных, сахар у астровых), белки (у фикуса), жиры танины, слизи, эфирные масла, каучук (более чем у 12 500 растений). Среди каучуконосов промышленное значение имеет тропическая гевея – Hevea (семейство Молочайные), в млечном соке которой содержится 40-50% каучука.
74. Стебель (у деревьев – ствол, ветви и побеги) служит связующим звеном между корнями, через которые вода и минеральные вещества поступают в растение, и листьями, в которых синтезируются питательные вещества. Проводящие ткани стебля составляют единое целое с аналогичными тканями корня и листа – это пути передвижения веществ. Клетки некоторых стеблей содержат хлорофилл, и в них происходит фотосинтез; другие клетки специализированы для запасания крахмала и иных питательных веществ. Стебель и ветви поддерживают листья в таком положении, чтобы на них падало возможно больше солнечного света, а цветки и плоды1–в положении, наиболее благоприятном для осуществления размножения. В точках роста стеблей образуются зачатки листьев и цветков. Стебли растений бывают травянистыми или деревянистыми. Мягкий, зеленый, довольно тонкий травянистый стебель типичен для однолетних растений. Такие растения образуются из семени, развиваются, цветут и дают семена в течение одного вегетационного периода, отмирая до наступления зимы. Другие травянистые растения являются двулетними – их жизненный цикл продолжается 2 года. В течение первого вегетационного периода в корне растения откладываются питательные вещества. Затем листья растения отмирают, а на следующий год из верхушки развиваются цветоносные стебли, которые дают семена. Примеры двулетников – морковь и свекла. Растения с мягкими и нежными стеблями, поддерживаемые преимущественно за счет тургорного давления, называются травами.
Стебли деревянистых растений в течение первого года жизни сходны по строению со стеблями травянистых. Однако к концу первого вегетационного периода в сердцевинных лучах возникает дополнительный камбий, так что в конечном счете он образует сплошное кольцо, проходящее как между сосудистыми пучками, так и внутри них. В течение каждого последующего года камбий продуцирует добавочные сдои ксилемы и флоэмы; эта флоэма замещает первичную флоэму и образует непосредственно снаружи от камбия тонкий непрерывный футляр из проводящей питательные вещества ткани, а ежегодные отложения ксилемы образуют годичные кольца. Побег (лат. córmus) — один из основных вегетативных органов высших растений, состоящий из стебля с расположенными на нём листьями и почками. Вегетативный невидоизменённый побег — единый орган растения, состоящий из стебля, листьев и почек, формирующийся из общего массива меристемы (конуса нарастания побега) и обладающие единой проводящей системой. Стебли и листья, являющиеся основными структурными элементами побега — часто рассматриваются как его составные органы, то есть органы второго порядка. Кроме того, обязательная принадлежность побега — почки. Главная внешняя черта, отличающая побег от корня — наличие листьев. Для побега характерна метамерность, то есть чередование одинаковых частей — метамеров. Листья располагаются на стебле в определённом порядке, в соответствии с типом листорасположения, а участок стебля на уровне отхождения листа называется узлом. Если основание листа полностью окружает стебель, узел называется закрытым, в противном случае — открытым. Промежутки стебля между
76. Понятие о филогенезе и онтогенезе . Органогенеза ФИЛОГЕНЕЗ (от греч. phylon - род - племя и ...генез), процесс исторического развития мира живых организмов как в целом, так и отдельных групп - видов, родов, семейств, отрядов (порядков), классов, типов (отделов), царств. Филогенез изучается в единстве и взаимообусловленности с индивидуальным развитием организмов - онтогенезом. Онтогенез индивидуальное развитие организма, совокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом от момента его зарождения до конца жизни. О. включает Рост, т. е. увеличение массы тела, его размеров, дифференцировку. (основные этапы индивидуального развития, возрастные изменения, вегетативный рост и репродуктивное развитие, взаимодействие с внешней средой) раскрывают многообразную картину жизнедеятельности растений, начиная от возникновения зародыша до отмирания растения, прошедшего весь свой жизненный цикл.
В этой общей сложной картине особенное значение имеет взаимодействие развивающегося растения с меняющимися условиями внешней среды и необычайно выраятенная способность растений к адаптации, совершенствованию приспособительных свойств к преодолению неблагоприятных условий для роста и развития.
Одним из наиболее ярких проявлений зависимости развития от условий внешней среды является фотопериодический контроль цветения длиннодиевных и короткодневных видов растений. Эта четкая зависимость, давшая в руки исследователей возможность строго контролировать развитие с помощью определенной длины дня, сделала короткодневные и длиниодневные виды растений наиболее удобным объектом для изучения сложных систем регуляции цветения. Органогенез (от греческого - орудие, орган и - рождение, происхождение) — совокупность процессов формирования и развития органов во время эмбрионального развития. Органогенез предшествует образование у зародыша зародышевых листков (экто-, энто- и мезодермы) в процессе гаструляции, протекающем в период с 7-го по 19-й день после оплодотворения яйцеклетки. Исходным моментом органогенеза следует считать обособление, основных зачатков будущих органов и тканей.
77.Образование и особенности флоэмы Флоэма (луб). Это сложная ткань, так как образована разнотипными клетками. Основные клетки проводящие, называются ситовидными элементами (рис.37). Проводящие элементы ксилемы образованы мёртвыми клетками, а у флоэмы они в течение периода функционирования сохраняют живой, хотя и сильно изменённый протопласт. По флоэме происходит отток пластических веществ от фотосинтезирующих органов. Способностью проводить органические вещества обладают все живые клетки растений. А отсюда, если ксилему можно обнаружить только у высших растений, то транспорт органических веществ между клетками осуществляется и у низших растений.
Ксилема и флоэма развиваются из апикальных меристем. На первом этапе в прокамбиальном тяже формируется протофлоэма. По мере роста окружающих тканей она растягивается, и, когда рост завершается, вместо протофлоэмы формируется метафлоэма. В стеблях большинства растений флоэма располагается снаружи по отношению к ксилеме, a листьях — обращена к нижней стороне жилок листовой пластинки, в проводящих пучках корней тяжи флоэмы и ксилемы чередуются. По своему происхождению флоэма делится на первичную, дифференциирующуюся из прокамбия и вторичную, дифференциирующуюся из камбия. Первичная флоэма, в свою очередь, подразделяется на протофлоэму и метафлоэму, в отличие от вторичной флоэмы, у первичной отсутствуют сердцевинные лучи. Клеточный состав и первичной, и вторичной флоэмы одинаков; они состоят из нескольких типов клеток различной морфологии, выполняющих различные функции: Ситовидные элементы (ситовидные клетки, ситовидные трубки и клетки-спутницы), обеспечивающие основной транспорт Склеренхимные элементы (склереиды и волокна), выполняющие опорную функцию Паренхимные элементы (паренхимные клетки), обеспечивающие ближний радиальный транспорт
80. Зона пустыни Пустыня — природная зона, характеризующаяся равнинной поверхностью, разреженностью или отсутствием флоры и специфической фауной.
Различают песчаные, каменистые, глинистые, солончаковые пустыни. Отдельно выделяют снежные пустыни (в Антарктиде и Арктике — арктическая пустыня). Самая известная песчаная пустыня — Сахара (самая большая песчаная пустыня по площади), занимающая всю северную часть африканского континента. Близки к пустыням полупустыни (опустыненные степи), также относящиеся к экстремальным ландшафтам. Всего пустыни занимают более 16,5 млн км² (без учёта Антарктиды), или около 11 % поверхности суши. С Антарктидой более 20 %.Пустыни распространены в умеренном поясе Северного полушария, субтропических и тропических поясах Северного и Южного полушарий. Характеризуются условиями увлажнения (годовая сумма осадков меньше 200 мм, в экстрааридных районах — менее 50 мм, а в некоторых пустынях осадков нет десятки лет; коэффициент увлажнения, отражающий соотношение осадков и испаряемости, — 0—0,15). В рельефе — сложное сочетание нагорий, мелкосопочника и островных гор со структурными пластовыми равнинами, древними речными долинами и замкнутыми озёрными впадинами. Эрозионный тип рельефообразования сильно ослаблен, широко распространены эоловые формы рельефа. Большей частью территория пустынь бессточна, иногда их пересекают транзитные реки (Сырдарья, Амударья, Нил, Хуанхэ и другие); много пересыхающих озёр и рек, часто меняющих свои очертания и размеры (Лобнор, Чад, Эйр), характерны периодически пересыхающие водотоки. Грунтовые воды часто минерализованы. Почвы развиты слабо, характеризуются преобладанием в почвенном растворе водно-растворимых солей над органическими веществами, обычны солевые коры. Растительный покров разрежен (расстояние между соседними растениями от нескольких десятков см до нескольких метров и более) и покрывает обычно менее 50 % поверхности почвы; в экстрааридных условиях практически отсутствует. Песчаные пустыни населены из растений в основном колючими кустарниками, из животных — пресмыкающимися и мелкими степными животными. В песчаных пустынях над местами залегания подземных вод встречаются оазисы — «островки» с густой растительностью и водоёмами. Снежные пустыни в основном находятся за полярными кругами и населены животными, устойчивыми к холоду.
82. Типы ветвления побегов. Типы ветвления У различных растений наблюдается несколько типов ветвления: дихотомическое, моноподиальное, симподиальное. При дихотомическом ветвлении конус нарастания расщепляется надвое. При моноподиальном ветвлении верхушечная почка функционирует в течение всей жизни, образуя главный побег (ось первого порядка), на котором в акропетальной последовательности развиваются оси второго порядка, на них — третьего порядка и т. д. При симподиальном ветвлении одна из верхних пазушных почек образует ось второго порядка, которая растёт в том же направлении, что и ось первого порядка, смещая в сторону её отмирающую часть. Впервые такое деление было предложено немецким ботаником Пфитцером в конце XIX века. Для Псилотовидных характерно дихотомическое ветвление Дихотомическое ветвление является наиболее примитивным типом ветвления, наблюдается как у низших растений (например у Bryophyta[1]), так и у некоторых высших сосудистых растений (например у Lycopodiophyta[2], некоторых Pteridophyta,[3]). При дихотомическом ветвлении конус нарастания делится надвое, вновь образующиеся апексы также делятся надвое и так далее. Различают так называемое изотомическое дихотомическое ветвление (образующиеся ветви равны по размеру) и анизотомическое (при котором образующиеся ветви неравны).[4] Моноподиальное ветвление Моноподиальное ветвление является следующим этапом эволюции ветвления побегов. У растений с моноподиальным типом строения побега верхушечная почка сохраняется на протяжении всей жизни побега. Моноподиальный тип ветвления часто встречается среди голосеменных растений[5], также встречается у многих покрытосеменных (например, у многих видов пальм, а также растений из семейства Орхидные — гастрохилусов, фаленопсисов и других). Некоторые из них имеют единственный вегетативный побег (например, Фаленопсис приятный). Моноподиальные растения — термин, наиболее часто используемый в описании растений тропической и субтропической флоры, а также в научно-популярной литературе по комнатному и оранжерейному цветоводству. Моноподиальные растения могут существенно различаться внешне. Среди них есть розеточные, с удлиненным побегом, кустовидные. Симподиальное ветвление У растений с симподиальным типом строения побега верхушечная почка, закончив развитие, отмирает или дает начало генеративному побегу. После цветения этот побег больше не растет, а у его основания начинает развиваться новый[6]. Структура побега у растений с симподиальным типом ветвления сложнее, чем у растений с моноподиальным типом; симподиальное ветвление является эволюционно более продвинутым типом ветвления.[источник не указан 1249 дней] Слово «симподиальный» образовано от др.-греч. - («вместе») и («нога»). Симподиальное ветвление характерно для многих покрытосеменных растений: например, для лип, ив[7] и многих орхидных.
84. Свет как экологический фактор Для зеленых автотрофных растений свет является одним из важнейших факторов жизни, поскольку представляет им необходимую лучистую энергию для фотосинтеза, т. е. участвует в образовании органических веществ, необходимых для роста и развития. Кроме того, свет оказывает непосредственное влияние на рост, на многие процессы дифференциации в клетках и тканях, на само органообразование. Для жизни растений важно, что в процессе фотосинтеза они продуцируют больше веществ, чем необходимо для покрытия расходов на дыхание, т. е. образуется положительный баланс веществ, без которого немыслим рост и существование растения Как и при каких условиях образуется положительный баланс веществ, эта проблема подлежит экологическому исследованию. Практиков сельского или лесного хозяйства интересует урожай, т. е. продуктивность самого фотосинтеза. А эколог должен изучить и понять причины различной продуктивности фитоценозов (в связи с различной интенсивностью света) в разных условиях. Кроме того, весьма важен вопрос, как распределяются ассимиляты, как они используются самим растением и в фитоценозе в целом, т. е. как свет влияет на продуктивность растительного покрова.
85. Соцветия . Классификация соцветий Соцветие (лат. inflorescentia) — часть системы побегов покрытосеменного растения, несущая цветки и в связи с этим разнообразно видоизменённая. Соцветия обычно более или менее четко отграничены от вегетативной части растения. биологический смысл возникновения соцветий — в возрастающей вероятности опыления цветков как анемофильных (то есть ветроопыляемых), так и энтомофильных (то есть насекомоопыляемых) растений. Закладываются соцветия внутри цветочных или смешанных почек. Классификация и характеристика соцветий[править] По наличию и характеру прицветных листьев (прицветников): Фрондозные (лат. frondis — листва, листья, зелень), или олиственные — соцветия, в которых прицветники имеют хорошо развитые пластинки (например, фуксия, фиалка трёхцветная, вербейник монетчатый). Брактеозные — соцветия, в которых прицветники представлены чешуевидными листьями верховой формации — брактеями (например, ландыш, сирень, вишня). Эбрактеозные, или голые — соцветия, в которых прицветники редуцированы (например, дикая редька, пастушья сумка и другие капустные (крестоцветные). По степени разветвления: Простые — соцветия, в которых на главной оси располагаются одиночные цветки и, таким образом, ветвление не превышает двух порядков (например, гиацинт, черёмуха, подорожник и др.). Сложные — соцветия, в которых на главной оси располагаются частные (парциальные) соцветия, то есть ветвление достигает трёх, четырёх и более порядков (например, сирень, бирючина, калина и др.). По типу нарастания и направлению раскрывания цветков: Рацемозные, или ботрические (от лат. racmus и греч. ботрион — кисть, гроздь) — соцветия, характеризующиеся моноподиальным типом нарастания осей и акропетальным (то есть направленным от основания оси к её верхушке) раскрыванием цветков (например, иван-чай, пастушья сумка и др.) Цимозные (от лат. cyma — полузонтик) — соцветия, характеризующиеся симподиальным типом нарастания осей и базипетальным (то есть направленным от верхушки оси к её основанию) раскрыванием цветков (например, медуница). По характеру поведения апикальных меристем: Закрытые, или определённые — соцветия, в которых апикальные (верхушечные) меристемы осей расходуются на образование верхушечного цветка (все цимозные соцветия, а также рацемозные некоторых растений: хохлаток, толстянок, колокольчиков и др.). Открытые, или неопределённые — соцветия, в которых апикальные меристемы осей остаются в вегетативном состоянии (ландыш, гиацинт, грушанка и др.).
86. Вегетативное размножение растений Вегетативное размножение - это размножение вегетативными органами растений - корнями, побегами или их частями. В его основе лежит способность растений к регенерации, к восстановлению целого организма из части. Усиление функции вегетативного размножения привело к значительному видоизменению органов. Специализированными побегами вегетативного размножения являются надземные и подземные столоны, корневища, клубни, луковицы и т. п. Органами вегетативного размножения могут быть и корни. У некоторых растений (осина, ольха, малина, калина, осот) на корнях закладываются придаточные почки, дающие начало придаточным побегам. При укоренении этих побегов и последующем отделении их от материнского растения появляются новые особи. Растения, на корнях которых образуются побеги из придаточных почек, называют корнеотпрысковыми, а побеги, развивающиеся из этих почек, - корневыми отпрысками. Слабее выражена способность к вегетативному размножению листьями. У сердечника лугового на зеленых листьях, расположенных в основании побега и прилегающих к влажному субстрату, закладываются придаточные почки. Прораcтание этих почек и укоренение вновь возникающих побегов обеспечивают вегетативное размножение растения. Совершающееся в природе вегетативное размножение растений называют естественным вегетативным размножением. Способность растения к размножению побегами и корнями издавна используется человеком в практике растениеводства. Искусственное вегетативное размножение растений обычно связано с хирургическим вмешательством и разделением целого организма на части. Вегетативное размножение широко используется человеком с целью получения урожая за более короткий срок и в больших количествах по сравнению с тем, что можно получить при размножении тех же растений семенами (например, размножение земляники столонами, картофеля - клубнями). Кроме того, вегетативно размножают растения тогда, когда необходимо сохранить сортовые качества сложных гибридов (от лат. гибрида – помесь), каковыми и является целый ряд разводимых и выращиваемых человеком растений. Возможно вообще отсутствие семян у бессемянных сортов. Такие растения размножают вегетативным путем.
88.Лесная зона . Под этим названием подразумевают более или менее значительное пространство, заросшее деревьями, и всю ту растительность, что покрывает почву между деревьями, состоя из кустарников, трав, папоротников, грибов и так далее. Эта растительность преимущественно может жить только под древесной тенью. Во времена доисторические, после ледникового периода леса занимали гораздо большее пространство, чем теперь. Хотя, по мнению некоторых ученых, периоду облесения местами предшествовал период степной. Большая часть Западной Европы, России и Сибирь представляла когда-то один сплошной лес. Вместе взятые леса земного шара все еще занимают приблизительно четверть поверхности, хотя эта оценка и не может считаться точной. Уменьшение леса шло рука об руку с увеличением народонаселения и развитием цивилизации, порождающей все более и более разнообразные потребности. В России, обладающей обширными тундрами и обширными солончаковыми, песчаными и глинистыми степями, тоже наблюдается сокращение лесных природных богатств. Всю страну, лежащую между 66° и даже 67° и 56° северной широты можно считать настоящим лесным царством. В этой полосе преобладает хвойный леса, или краснолесье, то есть сосна и ель. На востоке встречается лиственница, сибирская пихта и кедр. Также здесь произрастают: береза, осина, седая ольха, рябина, черемуха и некоторые высокоствольные ивы. В Сибири эта еловая или хвойная полоса простирается почти до 50° северной широты.
91.Отдел Диатомовые водоросли Диатомовые водоросли, диатомеи (от греч. diatomos — разделённый пополам) , кремнистые водоросли (Bacillariophyta), отдел (тип) водорослей (около 20 тыс. видов). Диатомовые водоросли микроскопические (0,75—1500 мкм), одноклеточные, одиночные или колониальные формы; среди последних встречаются; виды, живущие в слизистых трубках, образующие бурые кусты высотой до 20 см. Клетки диатомовых водорослей имеют твёрдый кремнёвый панцирь, состоящий из двух половинок, так называемых створок, находящих одна на другую. Верхнюю створку называют эпитекой, нижнюю — гипотекой. Стенки панциря имеют поры, через которые осуществляется обмен веществ с внешней средой. Многие диатомовые водоросли, у которых вдоль каждой половины панциря идёт щелевидное отверстие (так называемый шов), способны передвигаться по субстрату, видимо, за счёт выделения слизи. Клетки содержат одно ядро с одним или несколькими ядрышками и один или несколько хроматофоров жёлто-бурого цвета, из-за присутствия, наряду с хлорофиллом а, бурых пигментов (b-каротина и ксантофиллов); продукты ассимиляции — масло и волютин. Размножаются диатомовые водоросли делением; каждая дочерняя клетка получает половину материнского панциря, другая вырастает заново, при этом старая половина охватывает своими краями новую. Благодаря такому способу деления и тому, что пропитанные кремнезёмом твёрдые панцири мало или совсем неспособны к дальнейшему росту, диатомовые водоросли по мере размножения постепенно мельчают. При образовании ауксоспор (спор роста) содержимое клетки выходит из оболочки и значительно вырастает, давая начало новому, более крупному поколению . Ауксоспоры могут образовываться и половым путём, в результате слияния (конъюгации) содержимого двух клеток. У некоторых диатомовых водорослей наблюдаются размножение зооспорами и половой процесс с участием жгутиковых гамет (изогамия, гетерогамия или оогамия). У некоторых родов известны покоящиеся споры. Диатомовые водоросли диплоидны. Гаплоидны у них только гаметы.
94. Анатомическое строение стебля древесных растений на примере ветки липы. У липы сплошное камбиальное кольцо образуется в самом начале формирования стебля. Межпучковые зоны очень узкие, образуется сплошной цилиндр ксилемы (древесины) с очень тонкими прослойками основной ткани; во флоэме эти прослойки обычно шире, особенно в периферийной части. Благодаря деятельности сплошного кольца камбия первоначальный осевой цилиндр, свойственный первичному строению, разделяется на два слоя: внутренний – вторичную древесину и наружный – вторичную кору. Древесина – это ксилемная часть стебля, вторичная кора – флоэмная; вторичную флоэму называют так же вторичным лубом. В результате деятельности камбия в стебле накапливается все большая и большая масса элементов вторичного происхождения, а роль и значимость в общей массе первичных элементов структуры быстро сходит на нет. Следовательно, основная роль в формировании стебля двудольных, особенно древесных, принадлежит камбию. Формирование камбием элементов ксилемы и флоэмы происходит с неодинаковой быстротой; на одну клетку флоэмы камбий отделяет несколько клеток ксилемы; следовательно, ксилема нарастает гораздо быстрее, чем флоэма. У многолетних древесных растений к тому же происходит постепенное отмирание элементов флоэмы снаружи, поэтому в них на долю ксилемы приходится почти вся масса ствола и ветвей; флоэма же составляет сравнительно тонкий слой коры. Клетки камбия делятся преимущественно тангентально, благодаря чему эта ткань приобретает характерное строение: клетки располагаются правильными стопками, радиально направленными рядами. В связи с непрерывным утолщением ксилемного цилиндра окружность камбиального цилиндра также увеличивается, что происходит в результате радиального деления клеток камбия.За счет деятельности камбия в составе вторичных ксилемы и флоэмы формируются разнообразные элементы. Наибольшее разнообразие их достигается в многолетних стеблях древесных растений. Вследствие периодической деятельности камбия в ксилеме древесных растений возникают годичные кольца. Весной, благодаря обилию воды и питательных веществ, камбий формирует крупные элементы древесины; к концу вегетационного периода деятельность камбия затухает – в древесине всё больше преобладает механические элементы и узкие сосуды.
95. Гречишные, или Гречиховые (лат. Polygonáceae) — семейство двудольных растений, содержащее около 55 родов и более 1250 видов[3]. Сюда преимущественно относятся многолетние травы, хотя имеется и несколько древовидных и кустарниковых. Распространены почти повсюду, но больше всего в странах умеренного климата; в жарких странах травы этого семейства растут главным образом на горах; почти все древовидные гречишные растут в тропической Америке, а кустарниковые — в странах Средиземноморья. Листья у большинства цельные и очередные; сколько-нибудь раздробленные и даже сложные, так же как противоположные, составляют здесь редкость. Характерная черта семейства — наличие сросшихся прилистников — раструбов, иногда очень длинных. Цветки у большинства мелкие и неярко окрашены; околоцветник содержит от 4 до 6 частей, расположенных в 1 или 2 кружка; тычинок от 6 до 9, редко меньше или больше; свободная завязь, трёхгранная или сплюснутая, заканчивается тремя или двумя сильно развитыми столбиками; семяпочка одна, прямая, и прикреплена на дне завязи. Формула цветка:  [4], или
[4], или 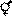
 В семействе Гречишных известно ветро- и насекомоопыление. В цветках гречишных насекомых привлекает нектар, который выделяют нектарники, расположенные у оснований тычинок, иногда в цветках имеются нектароносные диски. Опылителями являются насекомые с коротким хоботком, главным образом пчёлы и мухи. Плод сухой, односемянный, с плотно прилегающим к нему околоплодником, по большей части трёхгранный или сплюснутый; часто окружён околоцветником, иногда даже мясистым (у рода Muehlenbeckia Meissn.); только семя содержит мучнистую питательную ткань (перисперм) и хорошо развитый, различно искривлённый зародыш. Плоды Гречишных распространяются ветром, водой и грязью, прилипшей к ногам животных. Кроме семенного широко распространено и вегетативное размножение с помощью выводковых почек. Многие гречишные содержат обильные кислые соки и кристаллы щавелевокислой извести (щавель (Rumex)), а также смолистые, горькие вещества (ревень (Rheum)
В семействе Гречишных известно ветро- и насекомоопыление. В цветках гречишных насекомых привлекает нектар, который выделяют нектарники, расположенные у оснований тычинок, иногда в цветках имеются нектароносные диски. Опылителями являются насекомые с коротким хоботком, главным образом пчёлы и мухи. Плод сухой, односемянный, с плотно прилегающим к нему околоплодником, по большей части трёхгранный или сплюснутый; часто окружён околоцветником, иногда даже мясистым (у рода Muehlenbeckia Meissn.); только семя содержит мучнистую питательную ткань (перисперм) и хорошо развитый, различно искривлённый зародыш. Плоды Гречишных распространяются ветром, водой и грязью, прилипшей к ногам животных. Кроме семенного широко распространено и вегетативное размножение с помощью выводковых почек. Многие гречишные содержат обильные кислые соки и кристаллы щавелевокислой извести (щавель (Rumex)), а также смолистые, горькие вещества (ревень (Rheum)
96. Зона тундры. Тундра — вид природных зон, лежащих за северными пределами лесной растительности, пространства с вечномёрзлой почвой, не заливаемой морскими или речными водами. Тундра находится севернее зоны тайги. По характеру поверхности тундры бывают болотистые, торфянистые, каменистые. Для климата тундр характерна продолжительная и холодная зима, короткое лето, малое количество осадков, низкое испарение и высокая облачность. Тепловые условия определяются не только радиационным фактором, но и адвекцией, что особенно проявляется в европейской провинции тундр, где сказывается влияние Атлантического океана, Исландского барометрического минимума, но преобладает арктический воздух. Основной тип почв зоны тундрово-глеевый. Бедность органической массы вследствие медленного ее прироста, слабое разложение растительных остатков, малое количество микроорганизмов в почво-грунтах, особенно бактерий (азотофиксирующие бактерии почти отсутствуют), замедленность биохимических реакций, бедность минерального питания, доступного растениям, анаэробные условия - все это оказывает отрицательное влияние на активность почвообразовательного процесса.Наличие покрова из лишайников, мхов и торфянистого слоя чрезвычайно снижает снос мелких частиц в ручьи, реки и озера, поэтому средняя мутность речной воды снижена. Но коллоидная миграция значительна, вследствие чего быстро выносятся и минеральные коллоиды (гидроокислы железа и марганца). Речная и озерная вода, содержащая органические соединения, обычно имеет бурый цвет, относительно богата кислородом, что способствует обилию ихтиофауны. Флора тундр состоит из арктических и высокогорных, лесных и болотных форм умеренного пояса. Условия среды в тундре не благоприятствуют произрастанию сосудистых растений. Выживают лишь наименее требовательные бескорневые растения - водоросли, мхи, лишайники, имеющие широкое распространение, и отсутствуют клубневые. Вследствие короткого вегетационного периода растения-однолетники также почти отсутствуют, многолетники спешат закончить свой жизненный цикл, но не всегда это им удается. Некоторые растения выработали приспособления для защиты от излишнего света: блестящая листва, хорошо отражающая свет, особая пигментация и т. д. Интенсивное ультрафиолетовое излучение повышает содержание витаминов в ряде растений и обусловливает особенности их пигментации (интенсивная окраска цветов). Во время цветения воздух в тундрах насыщен фитонцидами. Среди цветковых преобладают растения с вечнозеленой листвой: брусника, голубика, багульник, водяника и др. Растения предельно используют тепло, не высоко поднимаются над почвой, образуют сланцевые и подушкообразные формы. Кустарники (ива), карликовая береза возвышаются над снегом и страдают от механических повреждений кусочками твердого снега, приносимого ветром. Часто можно наблюдать кустарники, как бы подстриженные на уровне снежного покрова.
97.Особенности образования и строения ксилемы. Её функция. Ксилема. В состав ксилемы входят: сосуды и трахеи, древесинные волокна и древесинная паренхима. Собственно проводящими элементами служат сосуды и трахеи. Они функционируют в отмершем состоянии. Древесинные волокна или либриформ относятся к механическим тканям, поэтому ксилема совмещает проводящие и механические функции. Паренхима служит в ксилеме запасающей живой тканью, способствующей связи ксилемы с окружающими ее живыми тканями. В многолетних частях растений древесинная паренхима может одревесневать.Сосуды (трахеи) – это глубоко специализированные элементы, представляющие собой длинные полые трубки. Они образуются из вертикального ряда прозенхимных меристематических клеток прокамбия или производных камбия. После увеличения этих клеток в длину и ширину продольные стенки их неравномерно утолщаются и одревесневают. Поперечные стенки перфорируются, т.е. в них образуются крупные отверстия. После этого протопласты отмирают, и весь вертикальный ряд клеток превращается в сплошную трубку. Участки сосуда, соответствующие бывшим отдельным клеткам, называются члениками. По форме и степени утолщения оболочек сосуды разделяются на несколько типов .Трахеиды существенно отличаются от сосудов тем, что представляют собой изолированные прозенхимные клетки, конечные стенки которых вытянуты и не перфорированы. Вода между соседнимитрахеидами передвигается через поры конечных стенок, поэтому скорость водного потока по ним значительно меньше, чем по сосудам. Из современных представителей растительного мира трахеидысвойственны высшим споровым и голосеменным растениям. У покрытосеменныхтрахеиды бывают в первичной ксилеме, во вторичной же развиваются преимущественно сосуды. Ксилема выполняет в растении две основные функции: по ней движется вода вместе с растворенными минеральными веществами и она служит опорой органам растения. Таким образом, ксилема играет в растении двоякую роль — физиологическую и структурную. В состав ксилемы входят гистологические элементы четырех типов: трахеиды, сосуды, паренхимные клетки и волокна.
98. Метаморфозы листа. Понятие об аналогичных и гомологичных органах. Видоизменения (метаморфозы) листьев— выработанные в ходе эволюции необратимые изменения формы листьев в результате приспособления органов растения к условиям среды обитания (т. е. с выполнением листьями новых функций). 1. Колючки — одно из наиболее часто распространенных видоизменений; они служат защитой от поедания животными. При этом лист либо целиком превращается в колючку (кактусы, молочаи, барбарис, белая акация, верблюжья колючка), либо в колючку превращается его часть (бодяк, чертополох, падуб). 2. Усики (у сложных листьев некоторых видов растений) цепляются за опору, вынося весь побег к свету. При этом в усик могут превращаться либо верхние листочки сложного листа (горох, вика), либо весь лист целиком, а функцию фотосинтеза выполняют прилистники (некоторые виды чины). 3. Запасающую функцию выполняют сочные чешуи луковиц (лук, чеснок), листья алоэ, кочана капусты. 4. Кроющие чешуи почек защищают нежные зачаточные листья и конус нарастания от неблагоприятных условий внешней среды. 5. Ловчие аппараты обеспечивают жизнь насекомоядных растений на болотах в условиях недостатка азота и других элементов минерального питания. Листья таких растений изменились до неузнаваемости, превратившись в ловушки (венерина мухоловка), кувшинчики (непентес). Листья некоторых растений своими блестящими, ярко окрашенными капельками на волосках привлекают муравьев, мух, комаров, других мелких насекомых; выделяющийся при этом сок содержит пищеварительные ферменты (росянка). Гомологичные органы - органы, сходные между собой по происхождению, строению, но выполняющие разные функции. Появление их — результат дивергенции. Пример гомологичных органов у растений: подземные корни растения, воздушные корни растений Разная среда обитания (разные условия) определяют появление гомологичных органов. заросток у простейших растений, эндосперм голосемянного растения, зародышевый мешок у покрытосемянных растений Это пример появления гомологичных органов в процессе эволюции (освоении суши). Аналогичные органы - органы и части животных или растений, сходные в известной мере по внешнему виду и выполняющие одинаковую функцию, но различные по строению и происхождению. Ключки на побегах (видоизменения побегов) и колючки - листья у голосемянных.
100.Биотические факторы. Биотические факторы – это совокупность влияний одних организмов на другие в процессе их жизнедеятельности (опыление растений, затенение верхними ярусами нижних, поедание одних особей другими). Влияние животных: 1.Пастьба животных оказывает существенное влияние на растительный покров пастбища. 2.Насекомые, реже птицы, опыляют растения. Отсутствие соответветствующих опылителей может быть причиной бесплодия растений. 3.Огромную роль играют представители животного мира и в процессе разноса плодов и семян. Такое распространение может быть и пассивным, и активным со стороны животных. 4.Велика экологическая роль землероев. Делая бесчисленные ходы в почве, и перерывая её, они могут улучшать ее воздушно-водный режим. В этом может сказаться положительная роль их. Вместе с тем, выворачивая на поверхность подпочву и образуя кочки, землерои отрицательно влияют на растительный покров, изменяя её состав в нежелательную сторону и ухудшая условия использования кормового угодья. 5.Общеизвестна положительная роль беспозвоночных, населяющих почву. Особенно велика роль земляных червей, отмеченная еще Ч.Дарвином. 6.Насекомые и их личинки нередко наносят огромный вред растениям, поедая корни или уничтожая листья. Взаимовлияние растений: 1.Произрастающие совместно растения затеняют друг друга. Поэтому в результате приспособления в естественных группировках сожительствуют растения с различной требовательностью к свету; под пологом более светолюбивых растений обитают растения тенелюбивые. Приспособления эти настолько глубокие, что после вырубки леса гибнет большая часть видов травянистых растений, обитавших под пологом деревьев, и возникает травянистая растительность иного состава. Деревья, выросшие в лесу, в окружении других деревьев, имеют совершенно иной вид по сравнению с деревьями, выросшими вне леса, на открытом месте. 2.Некоторые растения служат опорой для других растений, имеющих слабый стебель. Особенно распространено это в темных тропических лесах. 3.Совершенно односторонним и часто губительным для одной из сторон явл. Паразитизм растений на растениях. К числу наиболее известных растений-паразитов относятся многочисленные виды повилик и заразих. 4.Примерами обоюдополезного сожительства могут служить симбиоз бобовых растений и клубеньковых бактерий, корней высших растений и грибов ( микориза) и, наконец, тесный симбиоз между водорослями и грибами в едином организме лишайников. 5.Совместное обитание нескольких видов высших растений позволяет более экономно использовать факторы жизни растения: свет, влагу, элементы почвенного питания. На этом основаны преимущества возделывания смеси трав перед чистым посевами тех же видов.
101. Ботаника как наука, её задачи. Разделы ботаники. Ботаника – наука о растениях, изучает строение, жизнедеятельность растений в связи с условиями обитания биол. Наука, которая имеет большое значениеЗадачи ботаники: все стороннее изучение форм населяющих растений, распространение из по всему земному шару. Изучение внешних и внутренних их жизнедеятельности.изучение истор. Развития различных групп растенийизучение культурных растений с целью распространения и рационального использованияРазделы ботаники: анатомия растения, цитология, гистология (ткани), эмбриология, морфология (внешняя структура), физиология (обмен веществ, рост, развитие), хромография (определение и описание вида), систематика, география растений, экология ( условия обитания), микробиология, полеоботаника.
102. Система механических тканей. Механические ткани - это опорные ткани, придающие прочность органам растений. Они обеспечивают сопротивление статическим (сила тяжести) и динамическим (порывы ветра и т.п.) нагрузкам. Этим объясняется расположение тканей в органах растений, их тип и особенности клеток. В самых молодых участках растущих органов механических тканей нет, так как живые клетки в состоянии высокого тургора обусловливают их форму благодаря своим упругим оболочкам. По мере развития органов в них появляются специализированные механические ткани. Сочетаясь с другими тканями, они образуют как бы арматуру органа, поэтому их иногда называют арматурными. Механические ткани наиболее развиты в осевой части побега - стебле . Здесь они располагаются по его периферии: либо отдельными участками в гранях, либо сплошным цилиндром. Тем самым достигается наилучшее использование механических свойств ткани при изгибе органа. Напротив, в корне , который выдерживает главным образом сопротивление на разрыв, механическая ткань сосредоточена в центре. Механические ткани могут формироваться как в первичном, так и во вторичном теле растения. Наиболее заметная особенность клеток механических тканей - их значительно утолщенные оболочки, которые продолжают выполнять опорную функцию даже после отмирания их живого содержимого. Различают два основных типа механических тканей - колленхиму и склеренхиму
103. Степь в физической географии — равнина, поросшая травянистой растительностью, в умеренных и субтропических зонах северного и южного полушария. Характерной особенностью степей является практически полное отсутствие деревьев (не считая искусственных насаждений и лесополос вдоль водоёмов). Степи распространены на всех континентах, кроме Антарктиды. В Евразии наибольшие площади степей находятся на территории Российской Федерации, Казахстана, Украины и Монголии. В горах образует высотный пояс (горная степь); на равнинах — природную зону, расположенную между лесостепной зоной на севере и полупустынной зоной на юге. Атмосферные осадки от 250 до 450 мм в год. Средние температуры зимних месяцев от 0ºС до 20ºС, а летних от +20ºС до +28ºС. Климат степных регионов, как правило, находится в диапазоне от умеренно-континентального до резко-континентального, и всегда характеризуется жарким или очень жарким (до +40 °C) и сильно засушливым летом. Зима в степных регионах всегда малоснежная, с сильными позёмками и метелями, варьирует от умеренно мягкой до суровой с трескучими морозами, иногда возможны даже морозы до 40 °C. Характерная черта степи — безлесье огромных равнин, покрытых богатой травянистой растительностью. Травы, образующие сомкнутый или почти сомкнутый ковёр: ковыль, типчак, тонконог, мятлик, овсец и др. Растения приспосабливаются к неблагоприятным условиям. Многие из них засухоустойчивы либо активны весной, когда ещё остаётся влага после зимы. Так же степи богаты луковичными растениями (тюльпан) В зависимости от растительности и режима увлажнения степи подразделяют на пять основных подвидов: горные (криоксерофильные); луговые или разнотравные (мезоксерофильные) степи;настоящие (ксерофильные) с преобладанием многолетних дерновинных злаков, главным образом ковыля — так называемые ковыльные степи; сазовые (галоксерофильные) — степи, состоящие из растений, у которых надземные органы несут черты приспособления к засушливому климату, но произрастают при наличии постоянного или временного грунтового увлажнения; пустынные(суперксерофильные) степи с участием пустынных трав и полукустарников полыни и прутняка, а также эфемеров и эфемероидов. Также фрагменты отдельных типов степей имеются в лесостепи и в полупустыне. На разных континентах степь имеет разные названия: в Северной Америке — прерии; в Южной Америке — пампа, или пампасы, а в тропиках — льянос. Аналогом южноамериканских льянос в Африке и в Австралии является саванна. В Новой Зеландии степь называется туссоки.
106. Плод и его образование. Классификация плодов. Понятие о простом и сложном плоде. Соплодие. По мере развития семян завязь превращается в плод (настоящие плоды – костянка сливы, ягоды томата и смородины, боб гороха, стручки капусты), нередко в образовании плода участвуют и другие части цветка (ложные плоды): цветоложе (земляника), цветковые чешуи (злаки), цветоложе и чашечка (шиповник, яблоко). Преобразование целого соцветия образует соплодие (ананас, шелковица, инжир), при этом срастаются части цветков и главная ось соцветия.Функции плода – формирование, защита и распространение семян. Различают сухие и сочные плоды. Сухие могут быть вскрывающимися: листовка и многолистовка (магнолия, лютиковые), боб (бобовые), коробочка (маковые и лилейные), стручок (крестоцветные).Сухие невскрывающиеся: орех и орешек (лещина, земляника, шиповник), крылатка (ясень, вяз), семянка (подсолнечник и другие сложноцветные), зерновка (злаковые).Сочные плоды: костянка (вишня, слива), многокостянка (малина), ягода (смородина, баклажан, банан, томат), тыквина (огурец, арбуз, тыква), померанец, или гесперидий (апельсин, лимон).В распространении плодов принимают участие ветер, вода, животные и человек. Ягоды брусники, клюквы, смородины, костянки вишни, алычи, плоды рябины поедаются и разносятся птицами; желуди дуба, орехи лещины, сосны сибирский растаскиваются белками, мышами, кедровками, прячутся про запас в почву или трещины старых пней, где и прорастают. Плоды и семена бывают покрыты крючками и прицепками – цепляются к поверхности тела животных (семянки лопуха). С помощью ветра распространяются плоды с крыловидными выростами (клен, береза), хохолки, парашютики (одуванчик).Наружная часть плода – околоплодник (перикарп) – защищает от высыхания, механических повреждений, поедания (часто ядовитые, вяжущие, кислые вещества исчезают при созревании, либо обеспечивают избирательность поедания).В перикарпе выделяют эндокарп, мезокарп и экзокарп. В сухих плодах перикарп после созревания склерифицируется (клетки погибают).Плоды делятся на простые и сложные, или сборные. В тех случаях, когда в цветке имеется всего один пестик, из завязи его образуется после оплодотворения один плод, который называется простым (зерновка, боб, стручок и др.). Если плоды образуются не из одного, а из нескольких пестиков, они называются сборными, или сложными (сложная костянка у малины, ежевики, сложная листовка у лютика). В зависимости от строения околоплодника различают плоды сухие и сочные.Соплодие— совокупность зрелых плодов, образовавшихся в пределах одного соцветия, чётко обособленная от побега и других соплодий.Соплодия могут быть как достаточно рыхлыми, когда плоды сидят на обособленных плодоножках (виноград, рябина, калина и др.), так и более компактизированными — из сухих свободных односемянных плодов, окружённых обёрткой (астроцветные) или сочным вместилищем (инжир). При дальнейшей специализации соплодия срастаются голые плоды (панданус, ликвидамбар) или окружающие их околоцветники — сочные (шелковица) или сухие (свёкла). Крупные соплодия образуют ананасы, хлебное дерево, маклюры, при этом плоды срастаются с осью и другими элементами соцветия
. 109. Систематика механических тканей Общее свойство всех механических тканей – утолщённость клеточных оболочек. Утолщения могут быть равномерными и неравномерными. При всём значительном разнообразии, которое наблюдается в строении механических тканей, их можно разделить на три типа: колленхиму, склеренхиму и каменистые клетки, или склереиды. Каменистые клетки и клетки склеренхимы при окончательном оформлении ткани становятся мёртвыми; колленхима – живая ткань.Колленхима. Состоит из толстостенных клеток, способных к меристематической активности. В отличие от других механических тканей, оболочки клеток паренхимы кроме целлюлозы содержат большое количество пектина и гемицеллюлозы. Оболочки клеток начинают утолщаться на ранних этапах развития побега, но поскольку утолщение пластично и способно к растяжению, оно не препятствует удлинению стебля и листа. При возобновлении меристематической активности утолщения оболочки удаляются протопластом. В зависимости от характера утолщения оболочки различают: уголковую колленхиму (утолщение оболочки по углам), пластинчатую колленхиму (утолщение двух тангентальных оболочек) и рыхлую колленхиму (утолщение оболочки приурочено к межклетникам). Для колленхимы характерно периферическое положение в органах (стебле и листьях). Это прочная, гибкая ткань, состоящая из живых клеток с толстыми неодревесневшими оболочками.В старых частях растения колленхима может превращаться в склеренхиму – механическую ткань, имеющую равномерно утолщенные одревесневшие оболочки. В большинстве случаев это мертвая ткань. В зависимости от положения в органах растения и особенностей строения склеренхима представлена лубяными и древесинными волокнами. Последние отличаются сильно одревесневшими оболочками, короче лубяных, располагаются ближе к центру органа. Склереиды – каменистые клетки, встречающиеся в стебле, плодах, семенах, листьях. Они имеют разнообразную форму и встречаются по одиночке или группами, реже - в виде сплошного слоя (покровы скорлупы грецкого ореха, косточки сочных плодов, семенная кожура). Склереиды могут образовываться из клеток эпидермы, основной паренхимы и др. типов тканей.
112. Зонтичные (лат. Umbellíferae), или Сельдереевые[3], или Сельдерейные[3] (лат. Apiáceae) — семейство растений из порядка Зонтикоцветные (Apiales) класса Двудольные. Большинство зонтичных — многолетние травы, легко узнаются по характерным соцветиям — простые или сложные зонтики, цветки мелкие, большей частью белые, реже жёлтые или голубые, правильные, обоеполые, чашечка едва заметна, венчик с пятью лепестками, пестик один, завязь — полунижняя, двугнёздая. Формула цветка: [4]. 
 Плод — двураздельная семянка. Листья у большинства зонтичных перисто-рассечённые, часто с большим вздутым влагалищем.
Плод — двураздельная семянка. Листья у большинства зонтичных перисто-рассечённые, часто с большим вздутым влагалищем.
114. Образование и строение семязачатка. Типы семян. Образование и строение семязачатка. Развитие семязачатка начинается с закладки небольшого паренхимного бугорка внутри завязи пестика, разрастание его приводит к развитию центральной части – нуцеллуса. Покровы семязачатка (интегументы) закладываются в виде круговых валиков и, прорастая, со всех сторон облегают нуцеллус семязачатка. В верхней части они не смыкаются, оставляя щель (пыльцевход или микропиле). Противоположный полюс представляет собой основание и называется халазой. Здесь семязачаток прикрепляется посредством семяножки к стенкам завязи.Нуцеллус является мегаспорангием и вскоре после заложения семязачатка одна из его верхних клеток начинает разрастаться. Это археспориальная или материнская клетка мегаспор. Она делится мейотически и образует четыре гаплоидных мегаспоры. Три из них отмирают и только одна клетка сохраняется. Из нее и развивается женский гаметофит – зародышевй мешок. При этом мегаспора сильно разрастается, ядро ее претерпевает три последовательных митотических деления, в результате чего образуется восемь гаплоидных ядер, а остальные шесть расходятся к полюсам зародышевого мешка. На микропилярном полюсе формируется яйцеклетка и две синергиды, а на халазальном – три антиподы. Антиподы недолговечны и быстро расворяются, в некоторых случаях они выполняют роль сосущих образований – гаусторий. В такой фазе развития зародышевый мешок (женский гаметофит) готов к оплодотворению.Типы семян.Различают четыре типа семян в зависимости от места локализации запасных питательных веществ: с эндоспермом, с периспермом, с эндоспермом и периспермом и без эндосперма. Наиболее часто встречаются семена с эндоспермом и без него.Семя с эндоспермом характерно для многих однодольных и двудольных растений. Внутри семени формируется зародыш и эндосперм. Зародыш состоит из стебелька и зачаточнойпочечки, из которой при прорастании развивается надземная часть растения. Защитным органом почечки является колеоптиль. Он является наружным зародышевым листом. Частью зародыша является и щиток. Морфологически он представляет собой первый листочек зародыша и является единственно развитой семядолей. Физиологическая роль щитка сводится к всасыванию питательных веществ из эндосперма при прорастании семени. Редуцированной семядолей зародыша является эпибласт. Он находится на стебельке со стороны, противоположной щитку. Степень его развития влияет на засухоустойчивость растений. Подземная часть берет начало от зародышевого корешка, апикальная меристема которого прикрыта корневым чехликом. Вокруг зародышевого корешка имеется корневое влагалище колеориза. Колеориза выполняет защитную и всасывающую функции при прорастании корня. Большую часть семени занимает эндосперм с крахмальными зернами. В периферической его части, вдоль кожуры, расположен однородный слой клеток, содержащий белки (алейроновый слой).Таким образом, семя с эндоспермом состоит из семенной кожуры (спермодермы), зародыша и эндосперма.Семя без эндосперма состоит из двух частей – спермодермы и зародыша (фасоль, горох и т.д.). На семенной кожуре хорошо виден рубчик – это место прикрепления семени к семяножке. На одной линии с рубчиком, рядом с ним, расположено микропиле (через него осуществляется газообмен и водообмен). Над микропиле находится небольшой бугорок, образованный зародышевым корешком. С противоположной от микропиле стороны к рубчику примыкает семенной шов, это след от срастания семязачатка с семяножкой. Под спермодермой находится зародыш, состоящий из двух крупных семядолей почковидной формы, зародышевого корешка, зародышевого стебелька и почечки. Запасные питательные вещества откладываются в семядолях, клетки которых заполнены крахмальными зернами, расположенными в массе алейроновых зерен.