Фізичні властивості мембрани.
Тема: Біологічні мембрани
Вміти:
ü пояснити особливості структури біомембрани та її функції;
ü назвати і пояснити основні фізичні властивості мембран, які забезпечують життєдіяльність клітин;
ü охарактеризувати види та закономірності пасивного транспорту речовин;
ü пояснити роль осмотичного тиску плазми крові у перенесенні води;
ü пояснити роль онкотичного тиску в обміні водою між кров'ю і тканинною рідиною;
ü охарактеризувати механізм активного транспорту.
Вступ.
Основу всіх біологічних мембран складає подвійний шар молекул ліпідів, з якими зв'язані молекули білків. Незважаючи на те, що мембрани мають різний хімічний склад і виконують різні специфічні функції в різних клітинах, вони мають загалом універсальну будову і завжди виконують дві основні функції: слугують бар'єром для речовин та іонів і основою для мембранних ферментів, рецепторів та інших вмонтованих у мембрани білків.
Структура біологічних мембран.
Основу мембран становить біліпідний шар. У мембранах, що оточують клітину (плазматична мембрана), у цей шар входять також холестерин та гліколіпіди. На поверхні шару розміщені білки, які утримуються електростатичними силами і називаються периферичними. Інші білки контактують з гідрофобним шаром, з яким вони зв'язані гідрофобною взаємодією настільки міцно, що зруйнувати цей зв'язок можна, лише діючи на ліпідний подвійний шар органічними розчинниками. Такі білки називаються інтегральними. Загалом молекули фосфоліпідів є циліндричними, 1/3 або 1/4 з них — гідрофільні, а 2/3 або 3/4 — гідрофобні.
Білки, що входять до складу мембрани, становлять 70...75% її маси, а 25...30% — ліпіди. Біологічні мембрани порівнюють з ліпідним "морем", в якому плавають білки, причому море це бурхливе. Уся мембрана загалом — це динамічна рідкокристалічна система. Для рідкого кристала характерна, з одного боку, властивість рідини — плинність, а з іншого — певна впорядкованість в орієнтації молекул. Рідкі кристали — речовини, молекули яких мають форму планок, паличок, дисків, часто з чергуванням лінійних та циклічних угруповань атомів. Така форма молекул визначає паралельність їх розташування, тоді як у будь-якій рідині молекули розміщені хаотично. Останнім часом розвивається глобулярна теорія будови мембрани, згідно з якою мембрана
складається з ліпідних глобул, оточених білками і стабілізованих гідрофобною взаємодією поверхневого шару білкових молекул. Ці глобули мають форму правильних шести- або п'ятигранних призм, діаметром 8...14 нм. Форма глобул залежно від функціонального стану мембрани може змінюватись — ставати видовженою або стислою. Під час зміни форми глобул між ними можуть утворюватися пори, величина яких також змінюється. Перетворення глобул з однієї форми в іншу відбувається за рахунок енергії АТФ. Отже, мембрани можуть
мати два типи організації: пошаровий і глобулярний, які, можливо, переходять один в одний. Біологічна мембрана — це окрема фаза, що не змішується з водою. Ця фаза характеризується:
· поверхневим зарядом;
· поверхневим потенціалом;
· внутрішньою в'язкістю;
· здатністю розчиняти в собі або зв'язувати певні речовини;
· властивістю змінювати свій фазовий стан.
Мембрана може перебувати в одній з двох фаз, між якими відбуваються фазові переходи типу плавлення. За температури, меншої від температури плавлення, мембрана має властивості кристала, а за температури, більшої від температури плавлення, для неї характерна рідкокристалічна фаза, тобто це бімолекулярна рідка плівка.
Фізичні властивості мембрани.
Проникність мембрани для різних речовин, робота мембранних ферментів і рецепторів великою мірою зумовлені фізичними властивостями мембранної фази. Основною фізичною властивістю є наявність поверхневого заряду і зумовлена ним зміна електричного потенціалу на межі мембрана-водне середовище. Наявність заряду забезпечує стабільність мембранних структур, а також зв'язує неорганічні й органічні іони. Завдяки заряду на межі мембрана-вода виникає міжфазова зміна потенціалу того ж знака, що й заряд, а величина зміни потенціалу тим більша, чим більша поверхнева густина заряду. Зміна потенціалу впливає на здатність мембрани зв'язувати іони з навколишньої водної фази.
У середині, де концентрація іонів невелика, а розчинені в ліпідному шарі іони мало впливають на потенціал, мембрана за властивостями подібна до конденсатора. Потенціал поля конденсатора змінюється лінійно, а напруженість поля стала.
Один з методів визначення в'язкості мембран ґрунтується на вимірюванні поляризації флуоресцентного зонда, розчиненого в ліпідній фазі.
Суть методу у тому, що під час освітлення об'єкта поляризованим світлом флуоресценція теж буде поляризованою. Якщо за час свого життя в збудженому стані молекула не встигає обернутися, то інтенсивність поляризованого світла не змінюється ( Р0 ). Якщо ж молекула обертається — змінюється інтенсивність поляризованого світла. Молекули обертаються тим швидше, чим менша в'язкість. Цим методом можна визначити мікров'язкість ліпідів. Наприклад, під час підвищення концентрації холестерину в плазмі крові мікров'язкість ліпопротеїдів зростає. Це зумовлює порушення метаболізму ліпідів плазми.
Функції мембран
1. Фізичні та фізико-хімічні функції клітини полягають у хімічному метаболізмі і біосинтезі, біоенергетичних процесах накопичення енергії та її перетворення під час електро- і механічних процесів та транспорту молекул й іонів. Хімічна енергія АТФ перетворюється в хімічну, електричну, осмотичну і механічну роботу.
Біоенергетична й біосинтетична функції пов'язані між собою та із транспортуваннямречовин із середовища у клітину, і навпаки. Поєднання транспорту речовин зі збереженням вмісту клітини та її автономністю забезпечується клітинною мембраною. Клітинна мембрана – це напівпроникна перегородка завтовшки 7...10 мм, яка відіграє важливу роль в усіх функціях клітини. Мембрана має рідкокристалічну структуру, тому фізичні та хімічні закономірності рідких кристалів дають змогу зрозуміти механізм її функціонування. Досліджують роль рідких кристалів у виникненні деяких захворювань в організмі людини. Основна функція клітини - обмін
речовин. Рідкі кристали є ідеальним утворенням для цього.
Пружність мембран як рідких кристалів дає змогу пояснити явище фагоцитозу, що полягає у захопленні та поглинанні частинки мембраною. Внаслідок дотику до сторонньої частинки мембрана деформується, охоплює цю частинку і поглинає її. Таким чином частинка потрапляє в клітину.
Правильне розміщення елементів рідких кристалів — сприятливе середовище для дії внутрішньоклітинних каталізаторів. За певних умов рідкі кристали можуть набрякати, а потім стискатись. Форма рідких кристалів зручна для біологічних процесів. Вона поєднує в собі стійкість до зовнішніх дій з пластичністю, гнучкістю. Рідкі кристали надзвичайно чутливі до різних внутрішньоклітинних процесів, швидко реагують на них.
Вочевидь, відхилення від рідкокристалічного стану може зумовити порушення тих чи інших функцій, виникнення захворювання. Існує гіпотеза, що старіння організму тією чи іншою мірою пов'язане із кристалізацією мембран. Твердокристалічні ділянки клітинної мембрани втрачають рухливість, погіршують обмін речовин, і організм старіє. Внаслідок утворення злоякісних клітин також порушується рідкокристалічний стан мембрани. Доведено, що існує прямий зв'язок між вмістом холестерину в крові та ступенем розвитку атеросклерозу. До складу атеросклеротичних бляшок крім холестерину та його сполук входять фосфоліпіди, які відрізняють його від складу неушкодженої тканини. У бляшках ефірів вміст холестерину становить 95%, фосфоліпідів — 1%, а в неушкодженій тканині вміст ефірів — 39%, фосфоліпідів — 39%. Рідкокристалічна фаза зв'язує холестерин і перешкоджає утворенню ефірів холестерину. Внаслідок порушень надлишкові ефіри виділяються у вигляді твердокристалічних атеросклеротичних бляшок.
За допомогою мембрани вмонтовані у ній білки "дізнаються" про дію один одного. Наприклад, якщо один із білків змінить своє положення, то призведе до деформації мембрани, і пружна хвиля, яка поширюється мембраною, "сповістить" інші білки про те, що "відбулося" з білком. Деформація шару фосфоліпідів зумовлює поляризацію та виникнення електричного поля, яке полегшує транспорт іонів через мембрану.
2. Мембрани беруть участь у процесах обміну речовин, які забезпечують життєдіяльність клітин. В усіх мембранних структурах є ферментні системи. Результати досліджень свідчать, що поверхня мембран є тим місцем у клітині, де відбувається більшість біохімічних реакцій.
3. Мембрани координують і регулюють біохімічні і біофізичні процеси в клітинах. Вони сприймають зовнішні сигнали, перетворюють їх у команди, які регулюють обмін речовин усередині клітини. При цьому важливе значення має проникність мембран. Якщо змінюється проникність, то змінюються стаціонарні концентрації реагуючих речовин у клітинах, що спричиняє зміни швидкостей біохімічних і біофізичних процесів. Про це свідчить те, що біологічна дія більшості гормонів полягає у зміні проникності клітинних мембран. Під час порушення гормонального і вітамінного балансів в організмі порушується нормальний стан клітинних мембран. Виявлено збільшення проникності мембран лізосом і вихід у цитоплазму лізосомних ферментів при авітамінозі, гіпоксії, дії іонізуючого випромінювання. Такі ж препарати, як кортизон і гідрокортизон, здатні стабілізувати мембрани лізосом, що є однією з причин протизапальної дії цих сполук.
4. Клітинні мембрани забезпечують адгезію — зчеплення клітин під час утворення тканини. З'єднання клітин забезпечується наявністю на їхній поверхні специфічних структур у вигляді виступів і впадин. Цим зумовлюється механічне зчеплення типу "гніздо-шип", а також хімічне зчеплення. Міжклітинна рідина містить солі кальцію, які сполучаються з карбоксильними групами білків і фосфатними групами ліпідів, що зумовлює клейкоподібні властивості рідини. Стабільність тканин залежить від поверхневого заряду клітин. Тканини стабільні, якщо електричне відштовхування клітин зрівноважує сили їх молекулярного зчеплення. Проте клітини різних тканин мають неоднакову здатність до адгезії. Наприклад, мембрани клітин злоякісних пухлин втрачають структури, які забезпечують механічне та хімічне зчеплення. Вміст кальцію в пухлинах у 2 рази менший, ніж у нормі, а також зростають сили електричного відштовхування. Тому клітини пухлин є більш рухливими, вони відриваються від них, пливуть з течією рідини та утворюють метастази.
5. Мембрана визначає дихальну функцію клітин.
6. Мембрана — це середовище, в якому відбуваються біоенергетичні процеси: генерація біопотенціалів, окиснювальне фосфорилювання.
7. Мембрана бере участь у поширенні нервового імпульсу і визначає рецепцію (механічну, акустичну, смакову, зорову та ін.).
Мембранний транспорт.
Клітина — відкрита термодинамічна система, яка обмінюється речовиною, енергією й інформацією зі середовищем. Транспортні процеси, що відбуваються в мембрані, регулюють об'єм клітини і підтримують її іонний склад у вузьких межах коливань, а це створює необхідні умови для прояву активності ферментів. Вони зумовлюють іонні градієнти, потрібні для виникнення мембранного потенціалу, підтримання збудливості, а також транспортування деяких молекул та іонів. Системи транспорту в мембрані екстрагують зі середовища і концентрують у клітині речовини, що є джерелами енергії, а також необхідні для побудови компонентів клітини. Розрізняють такі типи транспорту:
· пасивний, зумовлений наявністю градієнтів;
· активний, пов'язаний з енергетичними затратами.
Біоелектричні потенціали
Вміти:
ü пояснити значення біоелектричних потенціалів для дослідження функціонування клітин;
ü назвати й охарактеризувати основні види потенціалів, які зумовлюють виникнення біоелектричних потенціалів;
ü записати формулу Нернста та проінтерпретувати її;
ü описати виникнення біопотенціалів клітин у стані спокою;
ü пояснити механізм генерації потенціалу дії;
ü охарактеризувати особливості поширення збудження нервовими волокнами.
Вступ
Електричні властивості виявляються під час функціонування нервової, м'язової, залозевої тканин і відображають фізіологічні процеси, що відбуваються в живому організмі. Електричні сигнали можуть бути локалізовані в певній частині організму або передаватись від однієї частини тіла до іншої.
Електрична активність — це властивість живої матерії, тому вивчення електричних явищ у живих об'єктах посідає важливе місце в біофізиці.
Види потенціалів.
Біоелектричні потенціали зумовлені потенціалами, що переважно виникають внаслідок нерівномірного розподілу іонів. Такими потенціалами є дифузійні, мембранні та фазові.
Дифузійні потенціали виникають на межі двох середовищ з різною рухливістю іонів або на межі двох розчинів одного й того ж електроліту з різною концентрацією.
Якщо в посудину з концентрованим розчином ZnSO4 обережно налити слабкий розчин тієї ж солі, то іони Zn2+ та SO4-2 почнуть дифундувати через межу розподілу і переносити електричні заряди. Однак швидкість дифузії іонів буде різною: 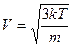
Маса іона цинку менша, ніж маса іона 50^ тому іони цинку випереджають іони кислотного залишку під час їх дифузії через межу як вгору, так і вниз. Проте концентрація нижнього розчину вища, і загалом знизу вгору буде перенесено більше іонів цинку, а на межі поділу виникне подвійний електричний шар. Додатний заряд зосередиться з боку менш концентрованого розчину. Між двома цими розчинами виникає дифузійна різниця потенціалів: 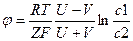
де U і V — відповідно рухливості катіонів та аніонів; с1 і с2 — концентрації розчинів.
Якщо ці два розчини розділені між собою мембраною, тобто поруватою перегородкою, проникною лише для іонів одного виду, то біопотенціали називають мембранними. Причиною виникнення таких біопотенціалів може бути різниця в діаметрах іонів, внаслідок чого великі іони не пройдуть через пори мембрани. Можливе також електричне відштовхування іонів одного знаку зарядженими кінцями дипольних молекул, що розміщені на поверхні мембран. Якщо, наприклад, мембрана не пропускає аніони, то рухливість їх під час дифузії можна вважати рівною нулеві, і тоді: 
У цьому випадку рівняння для мембранного потенціалу матиме вигляд: 
Це рівняння Нернста.
Мембранні потенціали можуть бути досить великими (50...55 мВ). Згідно зі сучасними уявленнями потенціали спокою, ушкодження, дії є за природою мембранними потенціалами.
Фазові потенціали виникають на межі двох фаз, які не змішуються. Такі потенціали виникають у клітинах на межі поділу різних фаз (компартмен-тів). За величиною вони незначні і визначаються також за формулою Нернста.
Потенціал спокою.
Основною причиною появи біоелектричних потенціалів є неоднакова проникність для різних іонів і виникнення внаслідок цього несиметричного розподілу іонів з обох боків мембрани.
Причини нерівномірності розподілу іонів та вплив на потенціал спокою можуть бути різними в різних клітинах.
Основні іони, які перебувають у великих кількостях у цитоплазмі клітини та в зовнішньому середовищі, — це іони К+, Na+, Cl- та іони великого діаметра, наприклад аніони амінокислоти R.
Нехай усередині клітини (В) є іони К+ і R-, а зовні (3) — лише іони К+,СІ-. Іони калію і хлору дифундують через мембрану в обох напрямах, а іон К. залишається всередині клітини.
Однак умови дифузії для іонів калію і хлору різні, тому що дифузії останніх зі зовнішнього середовища в клітину перешкоджає від'ємний заряд аніонів R-. Внаслідок цього концентрації іонів калію та хлору зовні і всередині клітини будуть неоднаковими.
Рівновага, що встановлюється в системі (рівновага Доннана), полягає в тому, що добуток концентрацій дифундуючих іонів з обох боків мембрани, тобто ззовні (3) і всередині (В), має бути однаковим: [К+]В[Cl-]В=[К+]З[Cl-]З, або [К+]В/[К+]З =[Cl-]В /[Cl-]З
Рівновага Доннана встановлюється між клітиною і навколишнім середовищем, якщо клітинна мембрана добре пропускає неорганічні іони, але не пропускає нуклеїнові кислоти та інші великі органічні іони. Рівновага Доннана найбільш характерна для мертвих клітин та клітин з послабленим метаболізмом. Для більшості клітин нерівноважний розподіл іонів створюється внаслідок активного транспорту іонів через мембрану транспортними АТФазами. Різниця концентрацій іонів усередині і ззовні клітин зумовлює виникнення потенціалу спокою на цитомембрані.
Різницю потенціалів між зовнішньою та внутрішньою поверхнями мембрани під час фізіологічного спокою клітини називають потенціалом спокою.
Аніони, для яких мембрана непроникна, концентруються біля внутрішньої її поверхні, створюючи від'ємний заряд, яким вони притягають зі зовнішнього середовища катіони калію, утворюють додатно заряджений шар на зовнішній поверхні мембрани. Наслідком цього є виникнення різниці потенціалів, яка визначається величиною мембранного потенціалу: 
де с1/с2— відношення концентрацій іонів калію.
Якщо температура клітини становить 20°С, а валентність іонів дорівнює одиниці, то, перевівши логарифм натуральний у десятковий, отримаємо:
Таким чином, наявність градієнта концентрацій іонів калію і хлору з обох боків мембрани призводить до виникнення біоелектричного потенціалу спокою, який є характерною особливістю живої клітини і становить близько 90 мВ.
Окрім калію, на виникнення потенціалів спокою впливають ще й інші іони. Наприклад,
різниця потенціалів, що виникає між ушкодженими та здоровими ділянками тканини, зумовлена градієнтом концентрації водневих іонів. Важливою є роль іонів Na+ У біологічних рідинах іонів Na+ є багато, але їхнє значення для виникнення біопотенціалів відрізняється від значення іонів К+. Усі живі клітини вміщують у 30 разів більше іонів калію, ніж їх є у міжклітинному середовищі, а іонів Na+ у клітині в 10 разів менше, ніж поза клітиною. Така різниця зумовлена різним ефективним діаметром іонів калію та натрію. Окрім цього, можна вважати, що, проникаючи всередину клітини, іони натрію виштовхуються з неї завдяки особливому механізмові, який називається натрієвою помпою. Проте невелика кількість іонів натрію все ж потрапляє у клітину, дещо зменшуючи потенціал спокою. Через клітинну мембрану дифундують також іони хлору, концентрація яких для більшості тканин вища в міжклітинному середовищі, ніж у клітині. Це також впливає на загальну величину мембранного потенціалу. Тому для точнішого визначення потенціалу використовують рівняння стаціонарного потенціалу Гольдмана-Ходжкіна-Каца:
де РK, РNа, РCl — коефіцієнти проникності для іонів калію, натрію та хлору; [К+], [Na+], [Сl+] - активності цих іонів всередині (В) та ззовні (3)
клітини.
З'ясовано, що в стані спокою проникність іонів К+ істотно більша, ніж Na+ і Сl+, а саме: для аксона кальмара РK : РNа : РCl = 1:0,04:0,45. Це підтверджує основну роль іонів К+ у генерації потенціалу спокою.
Швидкість зменшення мембранного потенціалу до критичного значення залежить від різниці потенціалу між збудженими й незбудженими ділянками і від кабельних властивостей волокна: ємності та опору мембран, ємності аксоплазми і середовища.
Залежно від типу клітин та умов їх перебування може переважати один із названих раніше механізмів виникнення мембранного потенціалу спокою, а саме:
· стаціонарний потенціал Гольдмана виникає внаслідок неоднакової концентрації катіонів і аніонів усередині і ззовні клітини, яка підтримується роботою мембранних помп. Рівняння Гольдмана точно описує потенціал спокою в нервових волокнах;
· потенціал, безпосередньо пов'язаний з роботою електрогенної помпи, виникає внаслідок активної роботи помпи і великого електричного опору мембрани, щоб струм, зумовлений активним транспортуванням, міг дорівнювати струму витоку за достатньо великого потенціалу. Цей механізм характерний для нервових клітин;
· якщо проникність мембрани для будь-якого іона істотно перевищує її проникність для інших, то виникає рівноважний потенціал Нернста, характерний для нервових і м'язових клітин і близький до калієвого потенціалу;
· доннанівський потенціал виникає, коли наявна висока концентрація непроникаючих іонів у клітині і висока проникність катіонів і аніонів, які входять до складу електроліту оточуючого середовища. Доннанівська рівновага іонів хлору створює потенціал на мембранах еритроцитів.
Потенціал дії.
У клітині внаслідок збудження виникає потенціал дії, який виявляється у швидкій зміні різниці потенціалів з обох боків клітинної мембрани. Потенціал дії спричиняє хвилю електричної активності, що поширюється нервовим або м'язовим волокном і відіграє роль переносника інформації або сигналу.
Причина цього явища полягає в тому, що дія на ділянку клітинної мембрани будь-яким чинником (механічним, тепловим, електричним) з силою, яка переважає певну межу, зумовлює відкриття натрієвих іонних каналів, і проникність цієї ділянки клітинної мембрани для іонів натрію зростає в сотні разів, тоді як для іонів калію — залишається незмінною. Внаслідок різниці концентрацій усередину клітини йде потік позитивно заряджених іонів натрію, які зменшують різницю потенціалів з обох боків мембрани. Це, у свою чергу, сприяє подальшому проникненню в клітину іонів натрію, які створюють на внутрішній поверхні мембрани, на ділянці дії чинника, позитивний заряд. Цей процес називається деполяризацією.
Поширення збудження
Поширення потенціалу дії зумовлене виникненням локальних струмів, які циркулюють між збудженою і незбудженою ділянками клітини.
У стані спокою на зовнішній поверхні мембрани є надлишковий позитивний заряд, а на внутрішній — негативний. Під час збудження відбувається перезарядження мембрани, внаслідок чого між збудженою і незбудженою ділянками поверхні мембрани виникає різниця і потенціалів. Вона зумовлює появу між цими ділянками локальних струмів, які йдуть на поверхню від незбудженої ділянки до збудженої, а всередині клітини — навпаки.
Локальний струм подразнює наступні незбуджені ділянки, спричиняючи збільшення проникності мембрани. Потенціал спокою зменшується. Коли деполяризація сягає критичного значення, на цих ділянках виникає потенціал дії, а на ділянках, які були збудженими в цей час, відбувається реполяризація.
Знову збуджена ділянка набуває від'ємного заряду; виникаючий локальний струм подразнює наступну ділянку. Цей процес повторюється, і таким чином збудження поширюється клітиною.
У нервовій системі імпульси поширюються в одному напрямі, оскільки синапси мають однобічну провідність.