В системній організації діяльності організму
Суми - 2009
Роль кори головного мозку
План лекції.
1. Структурно-функціональні особливості кори.
2. Роль кортикальних колонок.
3. Проекційні та асоціативні поля.
4. Методи вивчення кори великих півкуль.
5. Електроенцефалографія.
Кора великих півкуль з’являється вперше у риб у вигляді нюхових клітин, але видалення їх суттєво не впливає на поведінку риб. В амфібій вже є нюхова кора. Півкулі з’являються лише у птахів, у них при видаленні півкуль помітно змінюється поведінка: самі не можуть злетіти, самі не можуть їсти, в’ють гнізда, але не реагують на крик пташенят, у них зникає материнський інстинкт. Повного розвитку кора досягає у ссавців. У них видалення кори викликає втрату материнського інстинкту (собаки можуть поїдати своїх щенят), тварини не обходять перепони, натикаються на предмети, не йдуть на клич господаря, не орієнтуються в просторі, у них зникає захисний інстинкт. Найбільшу площу займає кора у приматів – 2200 см2. В них найкраще розвинені борозни та звивини. Функції кори повністю забезпечують пристосування до життя та вищу психічну діяльність. Видалення великих півкуль призводить до втрати здатності до самостійного життя.
У людини кора великих півкуль забезпечує такі функції:
- Взаємодія організму з навколишнім середовищем.
- Регуляція діяльності внутрішніх органів.
- Регуляція обміну речовин та енергії.
- Вища нервова діяльність –мова, пам’ять, мислення, свідомість.
Кора головного мозку є вищим відділом ЦНС. Це сіра речовина товщиною 3-5 мм, вкриває півкулі головного мозку. Вона займає площу 22 м2, утворюючи багаточисельні складки. В складі кори до 109-1010 нейронів, які утворюють 6 шарів:
1. Молекулярний шар – має мало клітин, їх волокна утворюють поверхневе густе тангенціальне сплетіння з дендритами другого шару.
2. Зовнішній зернистий шар – пірамідні клітини середньої величини, волокна яких розташовані радіально.
3. Внутрішній зернистий шар – зірчасті клітини, волокна яких розташовані горизонтально.
4. Внутрішній пірамідний (гангліозний) шар – це гігантські пірамідні клітини Беца, які мають довгі дендрити, що тягнуться до молекулярного шару.
5. Поліморфний шар – це шар веретеноподібних клітин.
Зв’язок кори великих півкуль з підкорковими структурами здійснюється за допомогою аферентних і еферентних волокон. Аферентні волокна називаються кортикопетальними, вони несуть інформацію в кору. Основними з них є таламокортикальн волокна. Це прямі аферентні шляхи, які розгалужуються у внутрішньому зернистому шарі і не дають колатералей. Невелика частина волокон йде в молекулярний шар, утворюючи колатеральні еферентні волокна, які називаються кортикофугальними, вони несуть інформацію від кори до підкоркових структур. Ці волокна діляться на 3 групи:
1. Проекційні – прямі еферентні волокна, що утворюють провідні шляхи (кортикоспінальні, кортикоталамічні)
2. Асоціативні - волокна, що утворюють безліч колатералей та йдуть в різні підкоркові зони однойменної півкулі.
3. Комісуральні - волокна, що йдуть від кори в складі мозолистого тіла і з’єднують зони кори однієї півкулі з підкорковими зонами другої.
1, 2 шари кори великих півкуль забезпечують аналіз та синтез отриманої інформації, мають багато асоціативних волокон.
3, 4 шари кори великих півкуль одержують інформацію від усіх органів та частин тіла за рахунок кортикопетальних волокон.
5, 6 шари кори великих півкуль – це рухові нейрони, звідси починаються рухові шляхи, що включають кортикофугальні волокна.
В шарах клітини розміщуються перпендикулярно до поверхні кори, утворюючи ланцюги. Елементарні нервові ланцюги відповідають за переробку певної інформації. Такий функціональний принцип названо – кортикальні колонки. Це елементарна функціональна одиниця, в якій здійснюється локальна переробка інформації від рецепторів однієї модальності. Кожна колонка має діаметр 500-1000 мкм, в складі яких розміщується 5-6 нейронів. Пірамідні клітини орієнтовані вертикально, їх аксони утворюють зворотні колатералі, які забезпечують як процеси полегшення в межах мікромодуля, так і гальмування між мікромодулями. Аксони зірчастих клітин ідуть через інтернейрони горизонтально, тому вони, головним чином, забезпечують гальмівні процеси. Веретеноподібні клітини мають довгі аксони, які орієнтовані як горизонтально, так і вертикально. Вони формують кортико-таламічні шляхи.
Мікромодулі об’єднуються в макромодулі завдяки горизонтальним розгалуженням терміналей. В колонці можуть бути прості та складні нейрони. Поряд з цим, в корі існує система, яка зчитує елементарні процеси в колонках та об’єднує всю інформацію. Формування таких систем зумовлено внутрішньо-кортикальними зв’язками між окремими макромодулями. Збудження одного мікромодуля викликає гальмування сусідніх. Активація мікромодулів відбувається за рахунок горизонтальних волокон таламокортикальних шляхів.
За функціями поля кори великих півкуль неоднозначні. Вивчення ролі окремих зон кори методом подразнення стало основою для вчення про локалізацію функцій в корі великих півкуль. Пізніше І.П. Павлов методом видалення в поєднанні з методом умовних рефлексів, підтвердив локалізацію функцій в корі. Разом з тим, Лешлі та ін. вважали, що різні зони кори великих півкуль є еквіпотенціальними, тобто однаковими за функціями – так виникла теорія еквіпотенціальності. Така думка була помилковою, бо тільки у нижчих тварин кора не має спеціалізованих зон. Так Лешлі, зруйнувавши кору щурів, знайшов, що порушення навиків руху щурів по лабіринту не залежить від місця руйнування. Досліди на тваринах з використанням мікроелектродного методу показали, що зони кори неоднозначні за функціями. Проекційні поля, створені мономодальними нейронами, одержують інформацію від рецепторів через переключаючі ядра таламуса. Вони оцінюють вид подразнення. На їх долю припадає 14% кори. Це специфічні проекційні поля. Асоціативні поля (86%) знаходяться навколо проекційних полів. Це полімодальні нейрони, які одержують інформацію від асоціативних ядер таламуса. Вони оцінюють джерело подразнення, його властивості та відстань до нього. Серед асоціативних полів є вищі поля, які забезпечують психічну діяльність людини, а також є поля, що формують пам’ять, індивідуальний досвід, прогнозування, передбачення. Це орбіто-фронтальні поля та тім’яна кора. Ці поля співпадають з функціональними зонами, де знаходяться нервові центри.
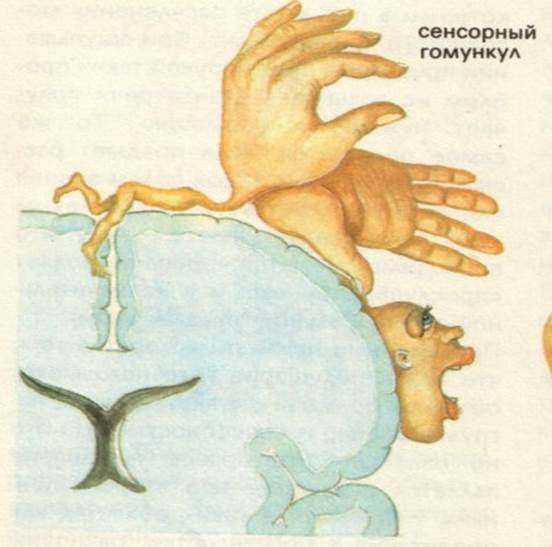
Представництво всіх видів чутливості в задній центральній звивині.
Задня центральна звивина – поля по Бродману – 1, 2, 3 – корковий центр всіх видів шкірної та пропріоцептивної чутливості. Вона має такі особливості:
1. Проекція протилежної сторони тіла розміщена вниз головою.
2. Має місце соматотопічна організація для кожної частини тіла.
3. Поля шкірної чутливості неоднозначні за площею, так, проекційні поля для обличчя, губ, кистей рук займають найбільшу площу.
При подразненні цієї зони виникає відчуття оніміння, тиску, повзання мурашок. При руйнуванні її зникає чутливість на протилежній стороні – анестезія.
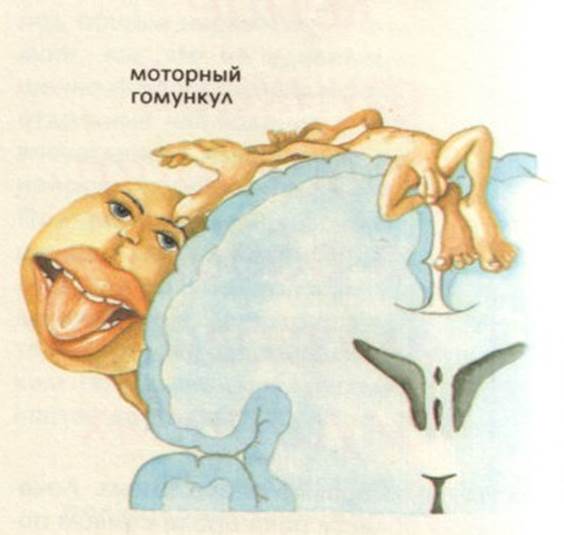
Проекція рухових центрів в передній центральній звивині.
Передня центральна звивина – поля по Бродману – 4, 6 – моторна зона. Вона забезпечує згинання, розгинання, пронацію, супінацію, ротацію. Вона також має соматотопічну організацію, але тут найбільшу площу займають мімічні м’язи обличчя, язик, кисті рук. При подразнення цієї зони виникає скорочення м’язів протилежної сторони тіла. При пошкодженні – розвиваються парези та паралічі. Парез – це обмеження довільних рухів. Параліч – це відсутність рухів.
Сенсорні системи мають подвійну організацію:
а) SS1 – це сомато-сенсорна зона – І, знаходиться в задній центральній звивині, виконує дискримінативний аналіз, тобто оцінює форму і характер поверхні предмету, приймає участь в складних рухах;
б) SS2 – це сомато-сенсорна зона – ІІ, має відношення до формування регуляції довільних рухів, орієнтації голови у напрямку звуків. Обидві зони мають сенсорні та моторні входи.
в) SМ1 – це сомато-моторна зона – І, знаходиться в передній центральній звивині;
г) SМ2 – це сомато-моторна зона – ІІ, знаходиться там же на медіальній поверхні кори. Обидві зони забезпечують відчуття просторової протяжності, стереогноз, відчуття ваги.
Потилична кора – це корковий центр зору. Він оцінює вигляд, форму, розміри та колір предмету. При пошкодження виникає геміанопсія (випадає частина поля зору), або виникає зорова агнозія (людина не впізнає знайомі предмети).
Верхня скронева звивина – корковий центр слуху (звивина Гешля – поля 41, 42). Оцінює звукові подразнення. При пошкодженні людина не може повторити сказане слово, хоч і чує, людина не розуміє звернену до неї мову, це слухова агнозія.
Задня частина нижньої лобної звивини (центр Брока) – це моторний центр мови. У правців він розміщується зліва. При пошкодження людина втрачає здатність до виразного мовлення, узгодження слів у реченні – моторна афазія.
Задня частина верхньої лобної звивини (центр Верніке) – це сенсорний центр мови, відповідає за сприйняття мови. При пошкодженні людина не розуміє звернену до неї мову – сенсорна афазія.
Задній відділ середньої лобної закрутки – руховий центр письма. При пошкодженні людина втрачає здатність писати – аграфія.
Кутова закрутка нижньої тім’яної долі (поле 30) – центр читання. При пошкодженні людина втрачає здатність читати – алексія.
Морський коник та його гачок – центр нюху. При пошкодженні людина втрачає здатність розрізняти запахи.
Верхні тім’яні долі – центр стереогнозу – оцінка положення тіла в просторі з закритими очима, глибока чутливість.
Надкрайова звивина(поле 40 лівої півкулі) – забезпечує здатність здійснювати привичні рухи чи автоматичні рухи, що сформувались в процесі життя. При пошкодженні розвивається моторна апраксія – людина знає, як виконати рух. але не може зробити його, не може намалювати предмет (порушується послідовність виконання складових руху).
Методи вивчення функцій кори великих півкуль:
1. Анатомо-клінічний метод – спостерігають за поведінкою людини при житті, вивчають структурні зміни після смерті.
2. Порівняльно-фізіологічний – оцінюють роль кори по поведінці у тварин різного рівня еволюційного розвитку.
3. Гістологічний – вивчають особливості будови кори на різних етапах розвитку організму.
4. Метод подразнення – вивчають функції кори шляхом подразнення різних її відділів.
5. Метод часткового та повного видалення кори – вивчають зміни поведінки тварин після видалення частин кори. (Роландо, 1823 рік).
6. Метод умовних рефлексів – І.П. Павлов вивчав поведінкові реакції у собак при сполученні різних подразників.
7. Електроенцефалографія.