Ретроингибирование и преодоление этого явления
Как можно масштабировать получение аминокислот в условиях биотехнологического производства?
Объясните процессы ретроингибирования, репрессии и их роль в получении конечного продукта.
Масштабирование – это увеличение объемов производства, в биотехнологии – переход от лабораторной установки к промышленному реактору.
Основной способ получения аминокислот - их микробиологический синтез основан на использовании биообъектов. В качестве биообъектов применяются штаммы-продуценты аминокислот – микроорганизмы рода Corynebacterium (почвенный микроорганизм), E. Coli (кишечная палочка — симбионт человека), Васillus subtilis (сенная палочка - почвенный микроорганизм).
Использование этих микроорганизмов для получения аминокислот основано на их способности самостоятельно синтезировать все 20 аминокислот.
Масштабировать производство аминокислот можно следующим образом
1) использовать мутантные штаммы-суперпродуценты
2) использовать оптимальную питательную среду для обеспечения высоких выходов аминокислот (концентрации источников углерода, аммонийного азота, минеральных солей, ростовых факторов, рН и температура).
3) Важный фактор, обеспечивающий высокие концентрации аминокислоты в культуральной жидкости – проницаемость клеточных мембран. Проницаемость клеточные мембран увеличивают либо с помощью мутаций, либо путем изменения состава питательной среды. Например, создают дефицит биотина (1-5 мкг/л) (биотин контролирует содержание в клеточной мембране фосфолипидов), добавляют пенициллин, детергенты или производные жирных кислот. Такие приемы повышают выделение аминокислот в среду и облегчают из последующую очистку.
Индукция и репрессия в биотехнологии
Образование многих первичных метаболитов (в том числе аминокислот) регулируется на нескольких уровнях.
1) конечные продукты метаболических путей (аминокислоты) ингибируют активность ферментов первых стадий биосинтеза (на уровне связывания с ферментами). Этот процесс называется ретроингибированием.
2) конечные продукты метаболических путей тормозят биосинтез ферментов последних его этапов (на уровне функционирования ДНК). Так низкомолекуляный метаболит может связаться с белком-репрессором, переводя его в активное состояние. Активный репрессор взаимодействует с определенным участком ДНК, блокируя связывание РНК-полимеразы. Этот феномен был назван репрессией.
Ретроингибирование и преодоление этого явления
Преодоление ретроингибирования проиллюстрируем на примере получения L-лизина.
Эффекта накопления в среде всего одной целевой аминокислоты добиваются путем блокирования процессов, ведущих к синтезу побочных аминокислот, возникающих в связи с разветвлением метаболического пути.
Образование лизина в клетке бактерии осуществляется под строгим метаболическим контролем. Аспартаткиназа открывает метаболический путь. Этот фермент ингибируется по принципу обратной связи при совместном и согласованном действии побочных продуктов L-треонина и L- 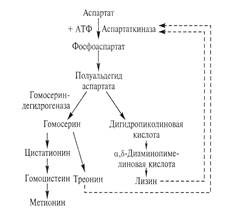
лизина.
При накоплении треонина и лизина в избыточной концентрации ингибируется аспартаткиназа и их синтез останавливается, при пониженной концентрации любой из двух аминокислот процесс активизируется.
Для получения лизина используются мутанты двух типов.
1) мутанты, у которых нет гомосериндегидрогеназы, в результате чего блокируется синтез метионина и треонина. У них существенно снижена внутриклеточная концентрация треонина, что снимает блокаду с аспартаткиназы. В среде, где присутствуют лимитирующие концентрации метионина и треонина, образуются избыточные количества лизина
2) Мутанты, дефектные по гену, определяющему конформацию аспартаткиназы. Такой мутантный фермент теряет чувствительность к высоким концентрациям аллостерического ингибитора — лизина.
Явление репрессии биосинтеза аминокислот рассмотрим на примере биосинтеза треонина. Как было показано, аспартаткиназа ингибируется по принципу обратной связи при совместном и согласованном действии треонина и лизина.
Когда одного механизма оказывается недостаточно и треонин продолжает накапливаться в избытке, у кишечной палочки включается еще один механизм регуляции биосинтеза - репрессия. При этом если треонин накапливается в избытке, он превращается в изолейцин, который, в свою очередь, также накапливается в избытке. В тот момент, когда треонин и изолейцин накапливаются в избытке одновременно, они опосредованно взаимодействуют с аттенуатором и подавляют транскрипцию, вызывая ее терминацию. В этом случае синтез всех белков-ферментов данного синтетического пути прекращается.