Полимеразная цепная реакция (ПЦР) - методика
Клонирование фрагментов ДНК. Общая схема.
клонирование ДНК: амплификация in vitro с помощью цепной полимеразной реакции или введение фрагмента ДНК в бактериальную клетку, которая после такой трансформации воспроизводит этот фрагмент в миллионах копий;
После того, как ДНК сшита в пробирке, ее необходимо размножить.
Существует два подхода к клонированию ДНК. Первый подход предполагает использование бактериальных или дрожжевых клеток для размножения введенной в них чужеродной ДНК. Второй способ представляет собой амплификацию ДНК in vitro.
Клонирование ДНК in vivo
Используя микроорганизмы, можно создавать два типа библиотек ДНК: геномную и клоновую (кДНК).
Геномная библиотека. Если геном какого-либо организма разрезать, вставить в плазмидные или вирусные вектора и ввести в клетку, то в таком виде его можно сохранить. При разрезании плазмидной или фаговой ДНК вероятность выпадения целых и неизмененных кусков генома довольно высока.
Такой способ получения геномной библиотеки получил название «метод дробовика», так как геном в данном случае представлен отдельными фрагментами.
Принципы создания плазмидных и вирусных векторов общие, поэтому рассмотрим их на примере плазмидных. Следует отметить, что из вирусных ДНК лучше использовать ДНК фагов, так как они имеют большую емкость и позволяют вставлять более крупные куски генома.
Очищенные кольцевые молекулы ДНК обрабатывают рестриктазой, получая линейную ДНК. Клеточную ДНК обрабатывают той же рестриктазой, добавляют к плазмидной, добавляют лигазы. Таким образом получают рекомбинантную плазмидную ДНК, которую вводят в бактериальные или дрожжевые клетки. Плазмида реплицируется с образованием многих копий. Многие плазмиды несут ген устойчивости к антибиотикам, и если в рекомбинантной плазмиде есть такой ген, то клетки легко выявлять, выращивая на среде с антибиотиком.
Каждая такая колония представляет собой клон или потомство одной клетки. Плазмиды одной колонии содержат клон геномной ДНК, а совокупность плазмид можно назвать библиотекой геномной ДНК. Недостаток такого метода в том, что фрагменты ДНК образуются в огромном количестве. Разрезание геномной ДНК определяется случаем, поэтому лишь часть фрагментов содержат полноценные гены. Некоторые фрагменты могут содержать только часть гена или же интронные последовательности.
Библиотека кДНК. Создание кДНК начинается с синтеза на матрице РНК с помощью обратной транскриптазы комплементарной нити ДНК. Затем создают щелочные условия, разрушают цепь РНК на нуклеотиды, после чего с помощью ДНК-полимеразы синтезируют комплементарную цепь ДНК. При этом образуется фрагмент ДНК с тупыми концами. Такую ДНК встраивают в плазмиды и вводят в клетки бактерий. При амплификации плазмиды образуется клон комплементарной копии ДНК (кДНК).
Преимущества клоновой ДНК перед клонами геномной ДНК в том, что кодирующая белок нуклеотидная последовательность гена ничем не прерывается.
Гены эукариот содержат интроны, которые должны удаляться из транскриптной РНК перед превращением ее в матричную, после чего следует сплайсинг (сращивание). Бактериальные клетки не могут осуществлять такую модификацию РНК, образовавшуюся путем транскрипции гена эукариотической клетки. Поэтому если преследуют получение белка путем экспрессии клонированного гена, то лучше использовать банк кДНК, полученной на основе матричной РНК.
Полимеразная цепная реакция (ПЦР) - методика
В 1985 году К. Мюллис с сотрудниками разработали метод клонирования последовательностей ДНК in vitro, который получил название полимеразной цепной реакции (ПЦР).
К анализируемому образцу ДНК добавляют в избытке 2 синтетических олигонуклеотида - праймера размером около 20 нуклеотидов. Каждый из них комплементарен одному из 3’-концов фрагмента ДНК. ДНК нагревают для разделения цепей двойной спирали, а при охлаждении происходит гибридизация праймеров с комплементарными участками фрагментов ДНК. В результате в растворе будут находиться однонитевые ДНК с короткими двухцепочечными участками - затравками (праймерами). При добавлении нуклеотидов и ДНК-полимеразы синтезируются комплементарные цепи и образуются идентичные фрагменты ДНК (первый цикл, рис. 43). Реакция останавливается и ДНК снова денатурируется прогреванием.
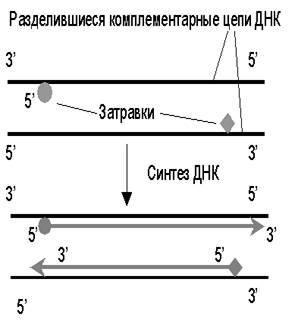
Рис. 43. Первый цикл полимеразной цепной реакции
В процессе охлаждения праймеры, находящиеся в избытке, вновь эффективно гибридизуются, но уже не только с цепями исходной ДНК, но и с вновь синтезированными. Внесение в систему ДНК-полимеразы инициирует второй цикл полимеразной реакции. Многократное повторение описанной процедуры позволяет провести 30 и более циклов ферментативного удлинения праймеров. При этом число сегментов ДНК, ограниченных с обоих концов используемыми праймерами, с каждым циклом ПЦР увеличивается экспоненциально (приближается к зависимости 2n, где n — число циклов). Выход всех других продуктов реакции увеличивается по линейной зависимости (рис. 44). Таким образом, в процессе рассматриваемой реакции эффективно амплифицируется только та последовательность ДНК, которая ограничена праймерами.
Первоначально для ПЦР использовали фрагмент Кленова ДНК-полимеразы I E. coli. Однако недостатком данного подхода являлось то, что после каждого цикла реакции необходимо было вносить в реакционную смесь новую порцию фермента. Кроме того, в оптимальных температурных условиях такой полимеразной реакции (37 °С) появлялись вторичные участки связывания праймеров и наблюдалась амплификация незапланированных сегментов генома, т. е. специфичность амплификации не была полной. Существенное улучшение метода полимеразной цепной реакции было достигнуто после замены фрагмента Кленова на ДНК-полимеразу термофильной бактерии Thermus aquaticus (Taq-полимераза). Температурный оптимум реакции, направляемой Taq-полимеразой, находится в районе 70 °С. Другим важным свойством является то, что данная полимераза не инактивируется после длительной инкубации при 95 °С.
Используя Taq-полимеразу, удалось решить сразу две проблемы. Во-первых, термостабильная полимераза не инактивируется на этапе денатурации ДНК, и поэтому нет необходимости после каждого цикла реакции добавлять новую порцию фермента. Такое упрощение процедуры позволило автоматизировать проведение ПЦР, так как теперь требовалось лишь перенесение образца с определенным интервалом времени в разные температурные условия: 90—95 °С (температура денатурации) и 60—70 °С (температура ренатурации ДНК и ферментативной реакции). Во-вторых, высокий температурный оптимум реакции, катализируемой Taq-полимеразой, позволяет подбирать жесткие температурные условия отжига, обеспечивающие гибридизацию праймеров только в заданном районе изучаемого генома, что существенно повышает специфичность и чувствительность метода.
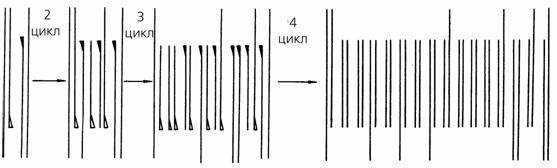
Рис. 44. Схема полимеразной цепной реакции
Используя метод ПЦР, можно in vitro селективно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион и более раз. Это позволяет надежно выявлять однокопийные гены и их варианты в таких больших и сложных геномах, каким является геном человека. Чувствительность метода такова, что амплифицировать в ПЦР и выявить целевую последовательность можно даже в том случае, если она встречается однажды в образце из 105 клеток. Получаемый сегмент ДНК надежно выявляется в виде дискретной полосы после электрофоретического разделения молекул ДНК и окраски их этидиум бромидом. Если к праймеру пришить фермент, то ферментная метка будет накапливаться при амплифицировании. Продукт амплификации проверяется по принципу ИФА, то есть добавляется субстрат и отмечается изменение окраски. В качестве метки можно использовать стрептавидин. Его можно пришивать как к праймеру, так и к нуклеотидам. В последнем случае нуклеотиды, меченные стрептавидином, добавляются к обычным, идущим на синтез комплементарной цепи ДНК. Этим достигается еще большее усиление сигнала.
Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. Поскольку при этом не требуется промежуточный этап клонирования фрагмента ДНК в молекулярных векторах, ПЦР иногда называют бесклеточным молекулярным клонированием (cell-free molecular cloning). Автоматизированная процедура Taq-полимеразной цепной реакции, состоящая из 30 и более циклов, занимает 3—4 часа, что существенно быстрее и проще процедуры клонирования определенного фрагмента ДНК в составе векторных молекул.
Применение полимеразной цепной реакции (ПЦР)
Используя метод ПЦР, можно in vitro селективно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион и более раз. Это позволяет надежно выявлять однокопийные гены и их варианты в таких больших и сложных геномах, каким является геном человека.
Чувствительность метода такова, что амплифицировать в ПЦР и выявить целевую последовательность можно даже в том случае, если она встречается однажды в образце из 105 клеток. Получаемый сегмент ДНК надежно выявляется в виде дискретной полосы после электрофоретического разделения молекул ДНК и окраски их этидиум бромидом. Если к праймеру пришить фермент, то ферментная метка будет накапливаться при амплифицировании. Продукт амплификации проверяется по принципу ИФА, то есть добавляется субстрат и отмечается изменение окраски. В качестве метки можно использовать стрептавидин (см. главу 3). Его можно пришивать как к праймеру, так и к нуклеотидам. В последнем случае нуклеотиды, меченные стрептавидином, добавляются к обычным, идущим на синтез комплементарной цепи ДНК. Этим достигается еще большее усиление сигнала.
Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. Поскольку при этом не требуется промежуточный этап клонирования фрагмента ДНК в молекулярных векторах, ПЦР иногда называют бесклеточным молекулярным клонированием (cell-free molecular cloning). Автоматизированная процедура Taq-полимеразной цепной реакции, состоящая из 30 и более циклов, занимает 3—4 часа, что существенно быстрее и проще процедуры клонирования определенного фрагмента ДНК в составе векторных молекул.
Полимеразную цепную реакцию используют для анализа индивидуальных вариаций последовательности нуклеотидов определенных локусов, для повышения эффективности клонирования целевых последовательностей изучаемых геномов и их прямого секвенирования, для детекции в организме патогенных микроорганизмов и т. п.
Используя 32Р-меченные синтетические олигонуклеотиды, можно выявлять единичные замены нуклеотидов в выбранных локусах геномной ДНК человека (или других организмов). Для этого в обычном варианте метода исследуемую ДНК гидролизуют рестриктазами, фракционируют электрофорезом, переносят разделенные фрагменты по Саузерну на нитроцеллюлозный фильтр, который гибридизуют с данным меченым олигонуклеотидом в условиях, при которых даже точечная замена нуклеотидов в анализируемой последовательности приводит к разрушению комплекса ДНК-олигонуклеотид.
Использование полимеразной цепной реакции для амплификации анализируемого локуса позволяет существенно упростить рассмотренный подход и повысить его чувствительность и специфичность. При этом для анализа аллельных вариантов генов достаточно всего 1 нг геномной ДНК человека, а гибридизацию можно проводить с негидролизованной рестриктазами ДНК, иммобилизованной на нитроцеллюлозном фильтре в виде небольшого пятна. Такой вариант метода позволил разработать новые диагностические тесты на генетические и инфекционные заболевания. В частности, этот подход используют для ранней диагностики наличия в организме вируса иммунодефицита человека (ВИЧ), что не удается осуществить другими методами. При этом не требуется работать с радиоактивными изотопами, так как амплифицированный сегмент вирусной ДНК выявляется напрямую после электрофоретического разделения ДНК и окраски их бромистым этидием.
Метод ПЦР позволил проанализировать наличие последовательностей вирусов папилломы человека в срезах биопсий новообразований шейки матки человека, залитых парафином за 40 лет до данного исследования. Более того, с помощью ПЦР удалось амплифицировать и клонировать фрагменты митохондриальной ДНК из ископаемых останков мозга человека возраста 7 тысяч лет!
На лизатах индивидуальных сперматозоидов человека продемонстрирована возможность одновременно анализировать два локуса, расположенных на разных негомологичных хромосомах. Такой подход обеспечивает уникальную возможность тонкого генетического анализа и изучения хромосомной рекомбинации, ДНК-полиморфизма и др. Метод анализа индивидуальных сперматозоидов сразу нашел практическое применение в судебной медицине, так как HLA-типирование гаплоидных клеток позволяет определять отцовство или выявлять преступника (комплекс HLA представляет собой набор генов главного комплекса гистосовместимости человека; локусы комплекса HLA — наиболее полиморфные из всех известных у высших позвоночных: в пределах вида в каждом локусе существует необычайно большое число разных аллелей — альтернативных форм одного и того же гена).
Используя ПЦР, можно выявлять правильность интеграции чужеродных генетических структур в заранее определенный район генома изучаемых клеток. Суммарная клеточная ДНК отжигается с двумя олигонуклеотидными затравками, одна из которых комплементарна участку хозяйской ДНК вблизи точки встраивания, а другая — последовательности интегрированного фрагмента в антипараллельной цепи ДНК. Полимеразная цепная реакция в случае неизмененной структуры хромосомной ДНК в предполагаемом месте встройки приводит к образованию фрагментов одноцепочечной ДНК неопределенного размера, а в случае запланированной встройки — двухцепочечных фрагментов ДНК известного размера, определяемого расстоянием между местами отжига двух праймеров. Причем степень амплификации анализируемого района генома в первом случае будет находиться в линейной зависимости от количества циклов, а во втором — в экспоненциальной. Экспоненциальное накопление в процессе ПЦР амплифицируемого фрагмента заранее известного размера позволяет визуально наблюдать его после электрофоретического фракционирования препарата ДНК и делать однозначное заключение о встройке чужеродной последовательности в заданный район хромосомной ДНК.