ПРОБЛЕМА КИСЛОРОДНОЙ ОПАСНОСТИ У РАСТЕНИЙ
Выше речь шла преимущественно о животных клетках. Однако проблема кислородной опасности стоит еще более остро у зеленых растений, клетки которых не только поглощают, но и образуют O2 . Быть может, именно защитой от кислорода объясняются многочисленные случаи так называемого несопряженного дыхания в растительных клетках.
Среди таких дыхательных систем, не способных к образованию ΔμH+ и, следовательно, к синтезу АТФ, наиболее изучена цианидрезистентная CoQH2-оксидаза. Фермент локализован во внутренней мембране митохондрий. Он катализирует четырехэлектронное восстановление О2 посредством CoQH2 . Интересно, что несопряженная CoQH2-оксидаза включается при гораздо больших отношениях CoQH2 / CoQ•-, чем сопряженная оксидаза, имеющаяся в тех же митохондриях. Напомним, что это отношение растет при переходе митохондрий в состояние покоя, когда возрастает риск "паразитных" реакций одноэлектронного восстановления О2 .
Quot;ДЫХАТЕЛЬНОЕ ПРЕДОХРАНЕНИЕ" У БАКТЕРИЙ-АЗОТФИКСАТОРОВ
Мысль о том, что дыхательная система может быть специализирована на снижении внутриклеточной [О2], первоначально обсуждалась применительно к двум проблемам, а именно биологической эволюции и фиксации азота. В первом случае предполагалось, что первичные дыхательные ферменты, возникшие в ответ на повышение количества О2 в атмосфере, имели своей функцией уборку О2 , продуцируемого фотосинтезом.
В клетках функция снижения уровня О2 , несомненно, присуща одной из дыхательных систем N2-восстанавливающих бактерий. Этот феномен был назван "дыхательным предохранением". Показано, что у таких бактерий поглощение О2 в процессе дыхания поддерживает концентрацию О2 на достаточно низком уровне, безопасном для нитрогеназы - фермента, восстанавливающего N2 и чувствительного к кислороду. Интересны результаты опытов на бактерии Azotobacter vinelandii. Эта бактерия характеризуется необычно высокой скоростью потребления кислорода. Дыхание сопровождается рассеянием столь больших порций энергии, что происходит быстрый разогрев ростовой среды. A. vinelandii обладает двумя конечными оксидазами типов o и d. Делеция в гене оксидазы o не влияет на аэробную фиксацию N2 , в то время как делеция в гене оксидазы d делает фиксацию N2 невозможной, если не понизить количество О2 до 1,5 объемных процентов. На основе этих фактов было сделано заключение, что оксидаза типа d ответственна за дыхательное предохранение у A. vinelandii при нормальном парциальном давлении кислорода.
Сосуществование нескольких конечных оксидаз в одной и той же клетке типично для многих видов бактерий. По-видимому, часть из них используется в качестве протонных насосов, часть - натриевых насосов, в то время как остальные катализируют несопряженное дыхание. Среди последних некоторые могут быть специализированы на "дыхательном предохранении", имеющем, вероятно, гораздо более общее значение, чем предохранение нитрогеназного механизма.
ЗАКЛЮЧЕНИЕ
Итак, кислород полезен для живой клетки как окислитель питательных веществ, но вреден как окислитель ДНК и других жизненно важных компонентов. Клетка располагает глубоко эшелонированной системой защиты от повреждающего действия кислорода. Эта система состоит из механизмов: (1) предотвращающих "паразитные" химические реакции одноэлектронного восстановления кислорода и (2) убирающих продукты такого восстановления.
Среди способов, предотвращающих зло, - уменьшение концентрации кислорода и его одноэлектронных восстановителей. Это может достигаться поглощением О2 , не сопряженным с синтезом АТФ вследствие (а) появления "неомической" протонной утечки в мембране митохондрий при повышении ΔμH+ над неким критическим уровнем; (б) образования пор в митохондриальной мембране; (в) активации особых дыхательных механизмов, не образующих ΔμH+ .
В борьбе с уже возникшими продуктами "паразитных" реакций кислорода участвуют: (1) супероксиддисмутаза, которая образует из не проникающего через мембрану O2•- проникающую Н2О2 ; (2) каталаза, разрушающая Н2О2 до О2 и Н2О; (3) пероксидаза, использующая Н2О2 для окисления определенных субстратов, (4) антиоксиданты типа витаминов Е, А и С, прерывающие цепные реакции, инициируемые продуктами "паразитных" реакций (рис. 4).
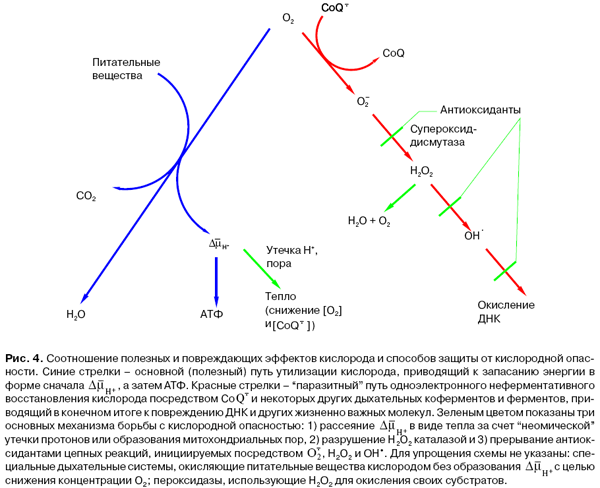
Клетки, не справившиеся с задачей защиты от кислородной опасности и тем самым поставившие под удар свой генетический аппарат, кончают самоубийством, включая апоптоз, зависящий от O2•-.
ЗАКЛЮЧЕНИЕ
Во всех клетках и всех их частях происходят образование АФК и ОМ макромолекул всех классов: нуклеиновых кислот, белков и липидов. Это минорные, но обязательные биологические процессы, выполняющие очень важные функции. Одна из них - синтез эйкозаноидных гормонов: простаноидов и лейкотриенов, а также участие в синтезе иодтиронинов. АФК и ОМ необходимы для иммунитета и воспаления, так как: а) увеличивают синтез цитокинов и иммунных рецепторов; б) способствуют выходу лейкоцитов в ткани; в) убивают фагоцитированные бактерии, старые и поврежденные клетки и способствуют повреждению несовместимых, а также злокачественных и пораженных вирусами клеток. Но избыточность АФК и ОМ повреждает клетки и может способствовать развитию очень многих болезней и синдромов, в том числе самых распространенных и опасных: старения, атеросклероза, инфаркта и инсульта, тяжелых воспалительных заболеваний, СПИДа, злокачественных процессов и др. Поэтому существует защитная антиоксидантная система. Она состоит из низкомолекулярных антиоксидантов и антиоксидантных ферментов. Соотношение прооксидантов и антиоксидантов и определяет, разовьется ли и будут ли прогрессировать оксидативный стресс и в результате свободно-радикальная патология. Это же соотношение участвует в регуляции многих очень важных биологических процессов, то есть существует новая форма контроля - редокс-регуляция..