Статева структура популяції. Третинне співвідношення статей. Фактори, які впливають на третинне співвідношення статей
Назвати типи стратегій поведінки популяції щодо зміни екологічних чинників (за Міркіним). Наведіть приклади.
Віолент (від лат. violent - нестримний, схильний до насилля) силовики "Лев", силовик .Види, що визначають вигляд угруповання, здатні до утискання конкурентів при відсутності порушень і за сприятливих умов. (трав’янисті рослини, чагарники, очерет, сосна у сосновому бору, лев, хижі тварини).
Патієнт (пристосуванці)"Верблюд" Види, здатні виживати в несприятливому середовищі при відсутності конкуренції. Тварини : верблюд, степова черепаха, варан, бобер, сліпак, олень-піщаник.
У рослин – види, які витримали вплив віолентів та підлаштувались під них. Рослини : білотка альпійська, аспленії, солонець європейський, осока колхідська, ранник крейдяний, гісоп крейдяний.
Експлерент (наповнювачі)"Шакал» Види, які швидко розмножуються і швидко розселюються в умовах дуже порушених біотопів. У тварин – види, які дуже швидко розмножуються. (крапчастий ховрах, миша домова, миша польова, заєць-русак) У рослин – види з низькою конкурентоздатністю, але властивістю швидко захоплювати звільнені території. ( лобода, щириця, лутига, осот польовий, полівки).
Статева структура популяції. Третинне співвідношення статей. Фактори, які впливають на третинне співвідношення статей.
Популяція – це мінімальна сукупність особин одного виду, які здатні до самовідтворення на протязі еволюційно тривалого часу, заселяють певний простір (ареал), утворюють самостійну генетичну систему і формують власну екологічну нішу.
Статева структура популяції – це числове співвідношення самців та самок популяції різних вікових груп. Виділяють 3 співвідношення статей: первинне – це числове співвідношення самців та самок на стадії зиготи, вторинне – при народженні, третинне – при настанні статевої зрілості.
Фактори, що порушують співвідношення статей: у самок лісового лемінга виявили самку з Х-хромосомою, в якій знаходиться фактор, що пригнічує дію Y-хромосоми. В результаті цього в популяції зустрічаються самки Х’Х (Х-фактор Х) ,
30% Х’ Y ху -
ХХ
Самці у дрозофіл з ознакою SR продукують Х-несучі сперматозоїди, тому майже все їх потомство несе Х-хромосоми з ознакою SR. Механізм цього явища полягає в елюмінації (виключенні) У – хромосомних сперміїв під час мейозу, під дією певного гена, який знаходиться в х – хромосомі. Все потомство буде складатися із самок.
Вплив факторів на вторинне і третинне співвідношення статей: варіює значно більше ніж первинне. Boniella viridis – самка впливає на ріст самців. Визначним є екологічні умови середовища. Самець паразитує на самці.
Arisxema japonica – вирішальним фактором при вторинному визначенні статі відіграє маса коренеплоду (розмір бульби) : якщо велика – жінка, статево розмножується; якщо мала – виростає чоловіча рослина і розмножується вегетативним способом.
Гормональне регулювання співвідношення статей : гормональна інверсія статі відбувається під впливом гормонів. В більшості хребетних доведено гормональне регулювання вторинного співвідношення статей. При розмноженні великої рогатої худоби відомі факти появи безплідних самок. Це явище вин при розвитку різностатевих близнюків ( чоловічі гормони пригнічають один із плоду, розвитку яєчників у самки) в результаті народжується самка, що не може завагітніти.
Гормональна інверсія статі – зміна статі після народження – це явище гермафродитизму розтягнутого в часі.
Протерандрія (протерандрія) – особина спочатку народжується як самець, а потім петворюється у самку, в такому випадку тварини є гермафродитами. (золотистий спар, або дорадо)
Явище протерогінія – особина спочатку народжується як самку, а з часом перетворюється в самця. Було описане у риб: морський карась, окунь, ставридових.
Атлантичний гігантський групер – спочатку як самець. При первинному нересті риба веде себе як самка, а в іншому нересті веде себе як самець – ангел-імператор.
Морський юнкер – спочатку самець, потім стає самкою, а потім знову стає самцем.
Стать залежить від температури. Гетерогонія – одна з форм вторинного чергування поколінь, при якій відбувається зміна статевих поколінь, які відрізняються одне від одного. Гетерогонія — видове пристосування для відтворення потомства у різних умовах розвитку. При паразитизмі гетерогонія дає можливість використати переваги існування всередині тіла хазяїна та забезпечує максимальне збільшення кількості народжуваних особин, тим самим сприяючи більшому поширенню даного виду.
Розрізняють чергування:
1) роздільностатевого покоління з гермафродитним (наприклад, у круглого черва рабдітіса (Rhabdonema nigrovenosum);
2) роздільностатевих поколінь, які розвиваються з запліднених яєць, з поколіннями, які розвиваються з яєць, що не потребують запліднення — партеногенетично (наприклад, у деяких попелиць);
3) статевих поколінь, які відрізняються за будовою (наприклад, деякі метелики).
Найпростішим випадком гетерогонії є розмноження тварини, при якому не відбувається чергування двох різних поколінь, але одна і та сама тварина спочатку відкладає протягом певного періоду яйця без участі самців, а згодом запліднені яйця.
Партеногенез або цикломорфоз.
Гетерогонія зустрічається серед різноманітних тварин: у плоских та круглих червів, комах та ракоподібних.
Прикладом чергування гермафродитних поколінь з роздільностатевими є круглий черв рабдітіс (Rhabdonema nigrovenosum). Гермафродитне покоління цієї тварини паразитує в легенях жаби. Молоді черви, які розвиваються з яєць, переходять до кишечнику жаби та разом з неперетравленими рештками потрапляють у вологу землю. Тут вони швидко розвиваються у самців і самок, які є роздільностатевим поколінням. З запліднених яєць після зараження жаби розвивається гермафродитне покоління.
Другий вид гетерогонії поширений серед комах. Філоксера (Phyloxera) являє собою ряд безкрилих поколінь, які розмножуються партеногенетично, потім у жарку пору року з'являються крилаті, теж партеногенетичні покоління, які відкладають яйця, з яких розвиваються самці та самки. Після запліднення самки відкладають одне велике зимуюче яйце, яке дає початок новому ряду партеногенетичних поколінь.
Класичним прикладом даного розмноження є попелиці (Aphidae). Протягом літа у них розвиваються ряд безкрилих або крилатих поколінь, які без участі запліднення народжують живих малят. Восени на світ з'являється роздільностатеве покоління, відбувається запліднення та відкладаються зимувальні яйця. При сприятливих умовах, наприклад, в теплицях, партеногенетичне розмноження попелиць може продовжуватися декілька років поспіль, не змінюючись розмноженням за участі обох статей.
Гетерогонія у найпростішому вигляді спостерігається у коловерток (Rotatoria) та дафнід. Незапліднені яйця, так звані літні, відкладаються у теплу пору року та мають ніжну оболонку; запліднені відкладаються восени при настанні несприятливих зовнішніх умов, вкриті товстою оболонкою та називаються зимовими. Нерідко перші та другі являють собою різкі відмінності у своєму розвитку.
Як показали дослідження, співвідношення числа самців і самок у популяції змінюється з часом, що зумовлене специфічною смертністю різних вікових груп та виживання потомства.
За останні роки отримані дані, які свідчать про можливість природного регулювання вторинного (у новонароджених) і третинного (у особин, що розмножуються) співвідношення статей внаслідок впливу несприятливих умов існування. Це є підтвердженням того, що в природних популяціях є приховані резерви регулювання структури, значення яких могло б стати основою при плануванні заходів захисту популяції.
У багатьох випадках антропогенний тиск скерований на одну із статей виду. Так, у птахів, копитних тварин, китоподібних внаслідок промислу найбільше скорочується кількість самців. Це суттєво порушує статеву структуру популяції.
Характеризуючи статеву структуру популяцій, необхідно звернути увагу на статевий добір, систему шлюбних відносин, статевий диморфізм. Вони впливають на розвиток популяції та її склад, оскільки відбирають сильніші і пристосованіші особини.
У співвідношенні статей важлива роль належить статевому диморфізму, тобто фізіологічним, морфологічним і поведінковим відмінностям статей. Він відіграє важливу роль у виборі партнера. Ті члени популяції, які знаходять найкращих партнерів для спаровування, в результаті дають краще потомство.
Статевий диморфізм іноді виконує й іншу екологічну функцію - зменшує конкурентну боротьбу між представниками різних статей за джерело їжі. Однією із переваг, даних індивіду статевим розмноженням, є те, що він може підвищити (або знизити) загальну пристосованість свого потомства, поєднавши свої гени з іншими генами (партнера), які впливають на виживання. Ця особливість має безпосереднє практичне значення і є основою селекційної роботи.
Типи росту чисельності популяцій. Адольф Кетле
Популяція будь-якого виду потенціально здатна до необмеженого зростання чисельності в ідеальних умовах. Швидкість її зростання за цих умов буде залежати лише від біотичного потенціалу особин даного виду. В природі, як уже наголошувалося, біотичний потенціал популяції повністю не реалізується.
Загальні зміни чисельності залежно від дії різних факторів відображені в чотирьох її типах: показниковий, логістичний, циклічний і стабільний.
Показниковий тип росту чисельності характеризується інтенсивним зростанням чисельності у геометричній прогресії (експоненціально) за умов відсутності лімітуючих факторів.
(Експотенційна або йод-подібна крива – це зміна чисельності за певний проміжок, яка відбувається по експоненті (поступове зростання). Ріст популяції в абсолютно оптимальних умовах (харчова база, відсутність конкуренції, відсутність сильного антропогенного фактору). Протягом короткого періоду часу.)
Однак, безмежне розмноження приховує в собі й велику небезпеку для будь-якої популяції, оскільки може призвести до швидкого підриву ресурсів середовища, нестачі їжі, сховищ, простору тощо. А це неминуче викличе загальне послаблення популяції. Перенаселеність несприятлива для будь-якого виду.
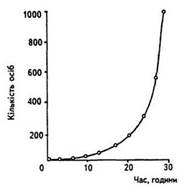 В основі цього типу лежить здатність організму надзвичайно швидко відтворювати собі подібних. Але для такого інтенсивного розмноження потрібні не тільки відповідний біотичний потенціал, а й відповідні оптимальні умови. Цей тип росту чисельності був відтворений у дослідах російського еколога Г.Гаузе (1945) в лабораторних умовах із туфелькою (Paramecium caudata), яка розмножується нестатевим способом (поділом на дві особини).
В основі цього типу лежить здатність організму надзвичайно швидко відтворювати собі подібних. Але для такого інтенсивного розмноження потрібні не тільки відповідний біотичний потенціал, а й відповідні оптимальні умови. Цей тип росту чисельності був відтворений у дослідах російського еколога Г.Гаузе (1945) в лабораторних умовах із туфелькою (Paramecium caudata), яка розмножується нестатевим способом (поділом на дві особини).
Такий експоненціальний ріст чисельності популяції продовжується до досягнення максимуму, після чого починають діяти лімітуючі фактори середовища і ріст популяції раптово припиняється. Цей тип росту чисельності популяції можна графічно описати J- подібною кривою. У природних умовах такий тип не спостерігається.
Приріст популяції пропорційний чисельності особин в ній, тобто
Де N –чи сельність популяції, а N – зміна чисельності за певний час.